
Full Text
The Roger Revelle Commemorative Lecture Series was created by the Ocean Studies Board of the National Academies in honor of Roger Revelle to highlight the important links between ocean sciences and public policy. Lisa A. Levin, the twentieth annual lecturer, spoke on April 24, 2019, at the National Academy of Sciences.
Why Focus on Deep Water?
The ocean covers 71% of our planet, but few people realize that most of the ocean consists of deep water, with an average depth of 3,800 m (nearly 2.4 miles). Because of their great depth, deep-ocean waters (below 200 m) represent over 95% of the volume on Earth that is habitable for animal life as we know it. This realm, largely out of sight and poorly explored, remains one of planet Earth’s final frontiers.
A Changing View of the Deep
Early imaginings of the ocean were as a home to sea monsters, like the kraken, a giant cephalopod in Norse mythology (Figure 1a). These monsters inspired fear, mystery, and some wonderful literature, such as Jules Verne’s 1870 classic 20,000 Leagues Under the Sea (Figure 1b). Despite the monster stories, the concept of the deep ocean as an azoic, lifeless desert dominated public perception until the 1860–1880s, when major expeditions (Lightning, Porcupine, Challenger) undertaking serious sampling of the deep sea provided consistent, incontrovertible evidence of an abundance of life in deep water (Koslow, 2007). The first hundred years of deep-sea exploration, conducted with sampling gear lowered from the ocean surface, painted a picture of a fairly homogeneous, mud-covered seafloor, understood to be cold, dark, environmentally stable, food-poor, and inhabited by small life forms. Sampling of deep-sea sediments in the 1950s and 1960s revealed these small inhabitants to be surprisingly diverse (Sanders and Hessler, 1969). Since that time, our understanding of the deep ocean and its ecosystems has changed enormously, enabled by technological advances that have allowed us to map the seabed at high resolution, generate startlingly detailed images in photographs and video, record animal sounds, document their mass migrations, even detect gas bubbles acoustically, and “sniff” the chemistry of seawater remotely. Today, use of a 4K camera at full zoom can effectively be like looking through a deep-sea underwater microscope. Remarkably, this viewing can occur not only in a remotely operated vehicle (ROV) control room aboard a ship but even in the comfort of one’s own home or office.
|
|

Using this array of tools, scientists have learned that the deep sea is not flat and homogeneous as depicted in early textbooks, but instead features tens of thousands of seamounts (underwater mountains) and continental margins punctuated by thousands of canyons. This steep topography supports gardens of filter-feeding and planktivorous sponges and corals that themselves provide habitat for a vast array of novel biodiversity. In the 1950s and 1960s, scientists found that potato-sized nodules high in manganese, nickel, cobalt, and other valuable metals cover vast areas of the abyssal plains at depths of 3,000–6,000 m, and interest grew in mining them as a source of metals (Mero, 1965). The discovery that hot fluids are emerging from Earth’s interior into the ocean along mid-ocean ridges and back-arc basins, and that they support chemosynthetic hydrothermal vent communities, occurred in the 1970s (Figure 2a). The next decade revealed that the subduction of oceanic plates under the continents creates methane-fueled gas seeps along continental margins, and that these seeps can also support chemosynthesis-based communities (Figure 2b). Today, we know of hundreds of vent and seep sites; but rather than a dichotomy, we are starting to recognize that there is a continuum of reducing, chemosynthetic ecosystems (Levin et al., 2012). Pelagic studies have gradually revealed that the deep ocean is a layer cake of distinct water masses and biogeochemical zones, formed by interactions with the atmosphere (wind), land, and seabed. Rather than being homogeneous, each depth region supports different communities of plankton, jellies, fish, and mammals (Sutton, 2013; Wisher et al., 2013; Choy et al., 2017).
|
|

Biodiversity Abounds
While only 5% of the seafloor has been studied and visualized by deep submergence vehicles, the last half-century of exploration has revealed a wealth of distinctive habitats, novel biodiversity, and unexpected animal lifestyles. In these newly discovered settings, tolerance to extreme conditions is a hallmark. Some vent animals can tolerate temperatures up to 50°C and a pH of 5, while microbes live at up to 120°C (McMullin et al., 2007). Halophiles can live at 20% to 30% NaCl (salt), seep biota tolerate hydrogen sulfide (a normally toxic chemical) at levels more than 1,000 times normal concentrations (>2 mM; Levin et al., 2013), fish can live at nearly zero oxygen concentrations (1 µMol kg–1; Gallo et al., 2018), and animals persist at pressures of nearly 1,100 atm in the bottom of the Mariana Trench
(Gallo et al., 2015).
Since the discovery of giant tubeworms living symbiotically on sulfide-oxidizing microbes, there has been continual discovery of novel symbioses between animals and bacteria that provide food to their hosts (Figure 3) and support high densities and sometimes very large body sizes (Dubilier et al., 2008). Structure-forming species such as cold-water corals and sponges can create massive reefs and provide attachment sites, nursery habitat, or spawning areas, and offer refuge from predators for myriad other species (Buhl-Mortensen et al., 2010), much like the aged trees in old growth forests. In fact, these areas have been called the rainforests of the deep because they represent a vast, long-lived reservoir of biodiversity, some of which enables them to survive changing environments through time. Scientists are finding this biodiversity is far more expansive and heterogeneous than we ever expected. Each seamount, canyon, seep, vent, ridge, and basin is slightly different.
|
|

A Cornucopia of Resources
The seemingly unending discovery of new biological diversity has uncovered a wealth of potential resources—both living and nonliving. Dense aggregations of bentho-pelagic fish have been discovered on seamounts, in canyons, and sometimes at methane seeps or on open slopes, fueling increasingly deeper fisheries (Morato et al., 2006; Watson and Morato, 2013). Orange roughy, Patagonian toothfish (Chilean sea bass), oreo, alfonsino, rockfish, thornyheads, and sablefish (black cod) are examples of fish species now harvested commercially from the deep ocean.
The seafloor is also a tremendous repository of minerals, trace metals, and rare earth elements. Their precipitation within polymetallic nodules across vast abyssal plains (Figure 4a; Cho et al., 2018), in polymetallic crusts on seamounts (Figure 4b; Yang et al. 2019), and as apatite in phosphorite nodules on continental margins (Figure 4c; Crosby and Bailey, 2012) is often mediated by microbes. Along with massive sulfides, precipitated at hydrothermal vents (Figure 4d) when hot metal fluids encounter cold deep-sea waters, these crusts and nodules represent very large potential sources of copper, cobalt, nickel, lithium, silver, gold, tellurium, rare earth elements, and phosphate (Levin et al., 2016; Cuyvers et al., 2018). Canyons and other slope environments sequester large amounts of dead organic material from the surface ocean that accumulates over geological time and ultimately, through the action of microbes, yields fossil fuel hydrocarbons in the form of oil, natural gas, and gas hydrates (frozen methane ice). Gas hydrates are found globally on continental margins and represent a vast untapped energy source of interest to some for mining. Thus, microbial diversity underpins the generation of non-living resources in the deep sea.
|
|

Less Tangible Ecosystem Services
We have also come to understand that the deep ocean provides a wealth of ecosystem services beyond the market services associated with fisheries, energy, and minerals, and that they, too, have value (Armstrong et al., 2012; Thurber et al., 2014; Le et al., 2017, Folkerson et al., 2018). Deep-sea biodiversity itself has value as a potential source of new pharmaceuticals, biomaterials, and other genetic resources (Blasiak et al., 2018). It provides key supporting services in the forms of food, habitat, and refuge (Buhl Mortensen et al., 2010), but also has inherent value, determined as willingness to pay to maintain species diversity (Ressurreição et al., 2011). Many believe that the deep sea functions as a living library of genetic adaptation capacity that will allow ecosystems and species to adapt to future conditions and allow humans to find solutions to future problems that have yet to emerge. The deep ocean also provides cultural services linked to scientific research, education, public viewing engagement, arts, and traditional values (Thurber et al., 2014). Many of the supporting services have only recently been discovered, for example, skate egg nurseries at hydrothermal vents (Salinas de Leon et al., 2018) and methane seeps (Treude et al., 2011; Sen et al., 2019), and fish eggs developing within giant protozoa called xenophyophores (author’s observation).
Regulating services provided by the deep ocean include large-scale cycling of nutrients and burial and sequestration of carbon (Armstrong et al., 2012; Thurber et al., 2014). The vast area of the ocean makes these services extensive and valuable. The biological pump, deep-sea animal bioturbation (Smith et al., 2008), and microbially mediated precipitation of carbonate at methane seeps (Marlow et al., 2014) all contribute critical carbon sequestration that reduces CO2 or CH4 concentrations in the atmosphere. New microbes discovered at hydrothermal vents and seeps have metabolic and enzymatic capabilities that make them potential agents of carbon removal from industrial emissions and the atmosphere (Mahon et al., 2015). Deep-sea microbes are being discovered that can break down plastic wastes, degrade natural and spilled oil, and remove other contaminants (Marietou et al., 2018; Teske, 2019).
Human Demands on the Deep Ocean
A growing human population, which has more than doubled in the last 50 years and is rapidly approaching 8 billion people, and higher household incomes have created rising demands for food, energy, raw materials, and telecommunications, as well as new pharmaceuticals and other genetic resources. As these resources become depleted on land, we look to the ocean, and increasingly into deeper waters, to supply our demands. This has led to a deepening of resource use across the fishing, energy, and telecommunication sectors, and the nascent development of deep seabed mining and deep bioprospecting industries (Merrie et al., 2014). In recent decades, new fisheries have developed for fish and shellfish from 400 m to 2,500 m depth. But there is a catch to this catch. Many deep-sea fish species are long-lived, with lifespans of 100 years or more. Without careful management of fish, fisheries, and their habitats, harvest of deep-sea fishes can be more akin to serial mining (Norse et al., 2012). The orange roughy fisheries have been depleted, seamount after seamount, although apparently sustainable populations have been identified (Doonan et al., 2015). Additionally, and perhaps more importantly, the harvest methods of bottom trawling and even longlining can destroy the three-dimensional structure created by habitat-forming species such as corals, sponges, and bryozoans (Clark et al., 2015, 2019).
Growing energy demands have led to increasingly deeper discovery and extraction of oil and gas, with wells now present to 3,500 m water depth (Merrie et al., 2014). Among the most famous of the deep oil wells is the Deepwater Horizon because of its blowout in the Gulf of Mexico in April 2010 and its generation of the industry’s largest ever oil spill. Gas hydrates are also being explored as an energy resource. Simultaneously, the industrial and transportation sectors have created a huge demand for steel, aluminum, and other metals. The rise of electronics (cell phones, computers) and green technologies in particular (solar panels, wind turbines, and hybrid car batteries) have accelerated the demand for less common metals such as copper, cobalt, nickel, zinc, and rare earth elements (Thompson et al., 2018).
The Growing Footprint of Human Activity
Human use of the deep ocean, and particularly exploration for and extraction of resources, creates physical, chemical, and biological disturbance. Introduction of sound and light, sediment resuspension, contaminant mobilization, hydrocarbon release, substrate removal, and alteration of the seabed geomorphology and geochemistry are all hallmarks of human presence; the effects can be far-reaching (Ramirez-Llodra et al., 2011; Merrie et al., 2014; Cordes et al., 2016; Levin et al., 2016). Continental margins, seamounts, canyons, and potentially nodule-covered abyssal plains are some of the settings that are, or will be, most heavily impacted.
Special features of deep-ocean ecosystems are likely to make them especially vulnerable to these disturbances. Many deep-sea organisms have life histories that confer great longevity—hundreds of years for fish (Norse et al., 2012) and thousands of years for some invertebrates such as corals and sponges (Roark et al., 2009; Jochum et al., 2017). These long-lived taxa are characterized by slow growth and maturation, and low reproductive and recruitment rates, and they can be rare; all of these traits can make recovery from loss or damage more difficult and in some cases improbable over human timeframes. The extremely high spatial heterogeneity now evident in many deep-sea settings, for example across seamounts and hydrothermal vents, means that species distributions are limited and even small-scale disturbance might risk biodiversity loss (Van Dover et al., 2017; Niner et al., 2018). A strong dependence of deep-sea species on specific hydrographic and geochemical environmental conditions suggests that physical disturbances that alter flow, substrate, fluid flux, or geochemistry will have profound effects on animal communities (Ramirez-Llodra et al., 2010; Levin et al., 2016; Gollner et al., 2017). Because deep-sea species exhibit highly developed biological interactions in the deep ocean, including endosymbiosis and numerous commensal relationships, changing conditions and loss of individuals or single species can impact many others.
Deep Ocean and Climate Change
The ocean is increasingly recognized as a mitigator of climate change. It absorbs about 93% of the excess heat in the atmosphere, and nearly half of this ends up in the deep ocean below 700 m (Glecker et al., 2016). The ocean has also removed about 30% of the excess CO2 generated by human activities since pre-industrial times (Gruber et al., 2019), and this too penetrates rapidly into deep water through overturning and thermohaline circulation (Gehlen et al., 2014). As it becomes a repository of heat and CO2, the deep ocean is becoming warmer, more acidic, and undersaturated with respect to carbonate. It is also losing oxygen, because of warming effects on solubility and vertical mixing (Levin and LeBris, 2015; Sweetman et al., 2017). These changes are accompanied by changes in stratification, nutrient availability, and surface primary production, which affect carbon export to the deep and food supply at depth. Vast areas of the seafloor beneath the oligotrophic ocean are predicted to experience reduced food supply, while the Arctic seabed is expected to receive more food (Jones et al., 2014).
These changes mean that areas currently supporting key species may not do so in the future, or may be able to support species not currently present. Although abyssal temperatures are understood to be highly stable, projected warmings of up to 1°C by 2100 (Sweetman et al., 2017) could yield habitat unsuitable if animals cannot adapt. The ocean has lost 2% of its oxygen over the last half century because of warming effects on solubility and stratification (Schmidtko et al., 2017), but this loss is greater at bathyal depths and can be up to 40% in some areas (Levin, 2018; Ren et al., 2018). Species have specific oxygen requirements and will migrate away or die if these are not met, and thus biodiversity loss is a major consequence of oxygen loss (Sperling et al., 2016).
Even at sublethal levels, oxygen loss may affect growth, reproduction, vision, disease resistance, and many other aspects of individual fitness (Breitburg et al., 2018). Some deep-sea animals are migratory and use multiple habitats during different ontogenetic life stages (i.e., larvae or juveniles) for reproduction and spawning; others move daily in diurnal vertical migration patterns. Warming or midwater oxygen loss may reduce the habitability of key migration corridors or lead to the shoaling of depth distributions. Expanding oxygen minimum zones, most evident at bathyal depths of 200–600 m (Stramma et al., 2010), will reduce the vertical migration depth of some zooplankton and fish (Bianchi et al., 2013; Netburn and Koslow, 2015), and compress the distributions of large billfish and tuna into shallower waters (Prince and Goodyear, 2007; Stramma et al., 2010; Mislan et al., 2017). Thus, ocean deoxygenation, which is often accompanied by high CO2 waters, is effectively a form of habitat loss for some taxa.
Will Climate Change Affect Recovery from Human Disturbance (or vice versa)?
The climate changes described above are superimposed on physical, chemical, and biological disturbance created by direct human activities (Figure 5). Both warming and ocean acidification increase metabolic rates and nutritional demands of animals; these effects will potentially create food shortages in already oligotrophic, food-poor environments such as abyssal plains. These settings are projected to experience further declines of up to 40% in food supply (particulate organic carbon flux to the seabed; Jones et al., 2014), creating additional stress on populations recovering in the direct footprint or plume shadow of
seabed mining.
|
|
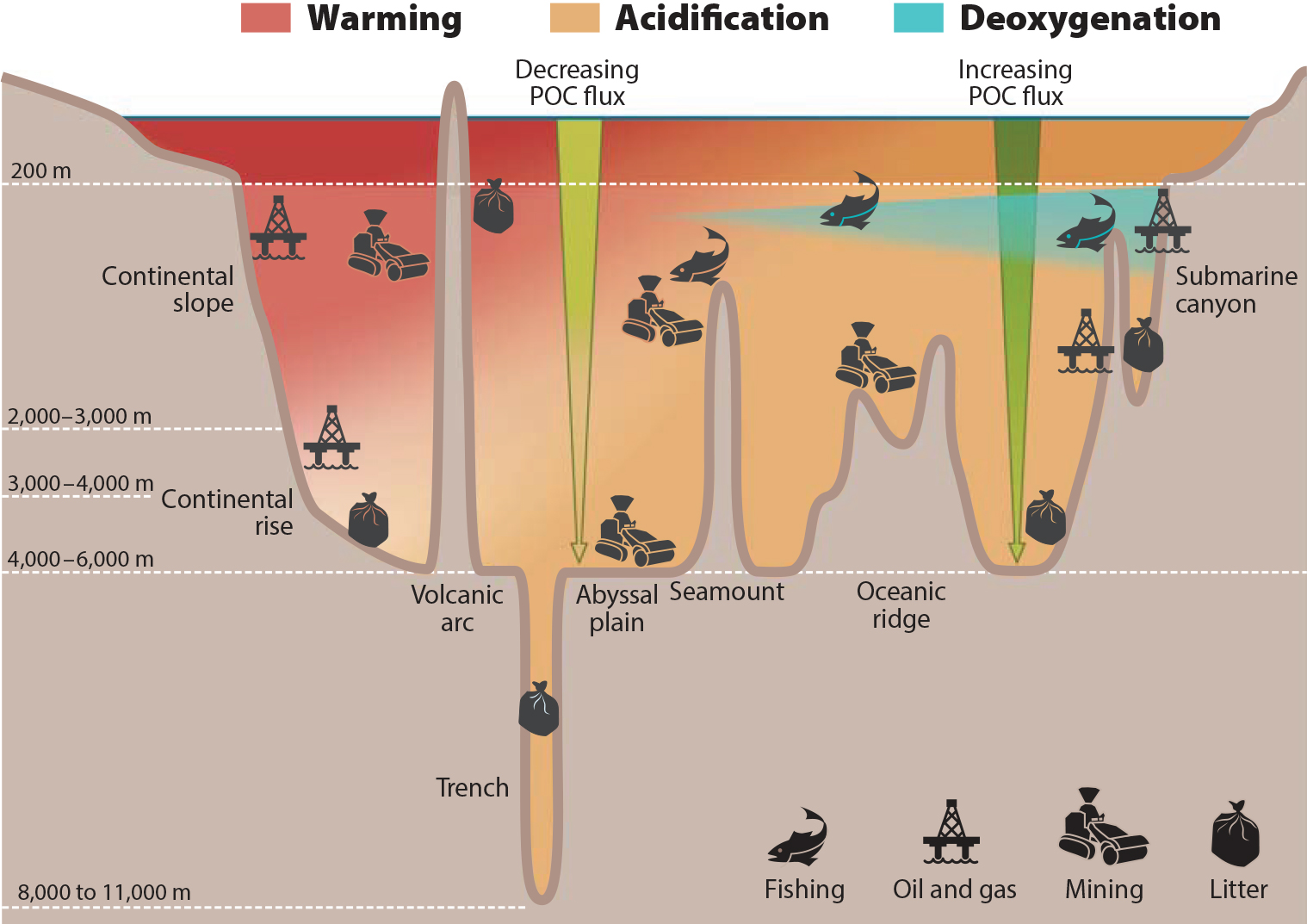
Projected declines in pH and carbonate saturation state can create conditions that are either unsuitable or energetically difficult for carbonate skeleton or shell formation, stressing deepwater corals, bivalves, brachiopods, and other calcifying taxa. Additionally, acidification will weaken and dissolve non-living portions of skeletons as well as the rubble that forms deepwater reefs, which are critical in supporting biodiversity (Hennige et al., 2015). Calcifiers damaged by trawling, anchors, mining, or other activities may recover more slowly in an acidifying ocean.
Whole assemblages subject to disturbance from sediment plumes, pollutants, or physical removal may become more vulnerable to oxygen stress, particularly in areas where oxygen is already low and climate-induced declines combine with direct disturbance (oil spills, plumes) to tip oxygen levels beyond thresholds. The frequency of different phases or regimes in major climate cycles may be changing in response to global warming. Interdecadal regimes such as the Pacific Decadal Oscillation and the North Atlantic Oscillation, and interannual cycles such as the El Niño-Southern Oscillation, can affect temperatures, oxygenation, and the carbonate system as well as circulation, eddies, and current strength and position. These in turn alter population connectivity, food supply, and habitat suitability (Fox et al., 2016). Thus, smaller body size and biomass, reduced productivity, loss of biodiversity, shifts in connectivity, and reduced resilience are some of the possible emergent properties resulting from cumulative impacts of climate change superimposed on direct human disturbance.
Managing for a Sustainable Future
As blue economies develop, and the human and climate footprints extend into the deep ocean, sustainable environmental management of deep-sea ecosystems and resources grows in importance. There are numerous uncertainties about how deep-sea ecosystems will respond to climate change and what this will mean for their resilience in the face of human activities. Proactive mitigation and adaptation responses are therefore justified. Ecosystem-based management has emerged as an essential approach for managing human activities over large ocean areas, including in the deep ocean. It is manifested in area-based management tools, in strategic environmental planning, development of guidelines for baseline data acquisition, impact assessment, and monitoring programs. As ecosystems entail communities interacting with their environment, it is evident that changing climatic conditions are a fundamental consideration for ecosystem-based approaches.
Climate Change and Protected Areas in the Deep Sea
One adaptation strategy for the conservation of species vulnerable to climate impacts is the protection of habitat that can serve as climate refugia (Morelli et al., 2016). Recognizing and delineating climate vulnerabilities can aid in area-based management to promote resilience by identifying sites least likely to change, those already degraded, and those destined to become newly suitable habitat. Areas considered less vulnerable to change, and with attributes soon to become scarce (such as high aragonite/calcite saturation state) could serve as valuable refugia. The Convention on Biological Diversity Subsidiary Body on Scientific, Technical and Technological Advice recommended, at Conference of the Parties 13, identifying and protecting areas known to be resilient to climate-related impacts and capable of acting as refugia sites in order to enhance the adaptive capacity of cold water ecosystems (UNEP/CBD/SBSTTA/20/4).
Efforts to build climate change into marine spatial planning have been strongest in the North Atlantic, partially because it is data-rich, and these data are accessible. Quieros et al. (2016) evaluated climate change and ocean acidification impacts in a spatially explicit model and co-mapped human activities in the Northeast Atlantic, generating recommendations about the long-term importance of protecting habitat refugia. Johnson et al. (2018) examined the efficacy of existing protected areas in the Northeast Atlantic under ocean acidification; this analysis revealed that many protected areas would be compromised by climate change over the next 20–50 years.
The International Seabed Authority is charged by the UN Convention on Law of the Sea with protection of the marine environment in areas targeted for deep-seabed mining. As part of this mandate, it has begun to identify, through regional environmental planning, areas of particular environmental interest (APEIs) where mining will not take place. The first consideration of climate change in the context of APEIs also concerned the North Atlantic, with year 2100 climate change projections at the seafloor incorporated into APEI scenario assessments for the northern Mid-Atlantic Ridge, the location of exploration claims by France, Russia, and Poland for mining of seafloor massive sulfides (Dunn et al., 2018; Figure 6). However, there are difficulties in applying climate projections without detailed information about the distributions and physiological thresholds and vulnerabilities of key taxa present.
|
|

The identification of Ecologically and Biologically Significant Areas (EBSAs) by the Convention on Biological Diversity (CBD) was carried out largely without climate considerations, but there is a recent proposal that climate change refugium be added as an eighth EBSA criterion applicable to all open-ocean and deep-sea habitats (Johnson and Kenchington, 2019). One of 300 existing EBSAs, the Western South Pacific High Aragonite Saturation State Zone, was developed to address climate change; it was identified as having the highest aragonite saturation state recorded in the Pacific and thus may be the area least affected by ocean acidification, conferring uniqueness and rarity (Johnson et al., 2018). EBSAs may become a starting point for biodiversity protections within ongoing (Intergovernmental Conference on Marine Biodiversity of Areas Beyond National Jurisdiction, or BBNJ) treaty negotiations. Building climate change into the planning can improve effectiveness of area-based management.
Another arena of deep-sea activity likely to be affected by changing conditions is fisheries. The management of bottom fisheries in the high seas (beyond national jurisdiction) in most regions is under the control of Regional Fisheries Management Organizations (RFMOs) within the Food and Agriculture Organization of the United Nations. They identify vulnerable marine ecosystems (VMEs), which become closed to fishing and require environmental impact assessments (EIAs) for new fisheries or fishing grounds. In December 2016 the UN General Assembly passed a resolution that called upon states and RFMOs to take into account the potential impacts of climate change in managing deep-sea fisheries and protecting vulnerable marine ecosystems. A subsequent technical report (FAO, 2019) combines climate projections, cumulative hazard assessment (Figure 7), climate vulnerabilities, and habitat suitability modeling to examine climate effects on deep-sea habitats, fish, and fisheries at depths between 500 m and 2,500 m, where most deep bottom fishing occurs. The North Atlantic and the Southern Ocean are identified as among the most susceptible to changing distributions of demersal fish and VME indicator species such as corals and sponges. Recommendations include integrating climate considerations into impact assessment, engagement of the industry for monitoring and mapping of bottom fishing areas and response to climate change, increased use of sensors for adaptive management, and more effective avoidance of VMEs and bycatch to reduce cumulative impacts from climate change.
|
|
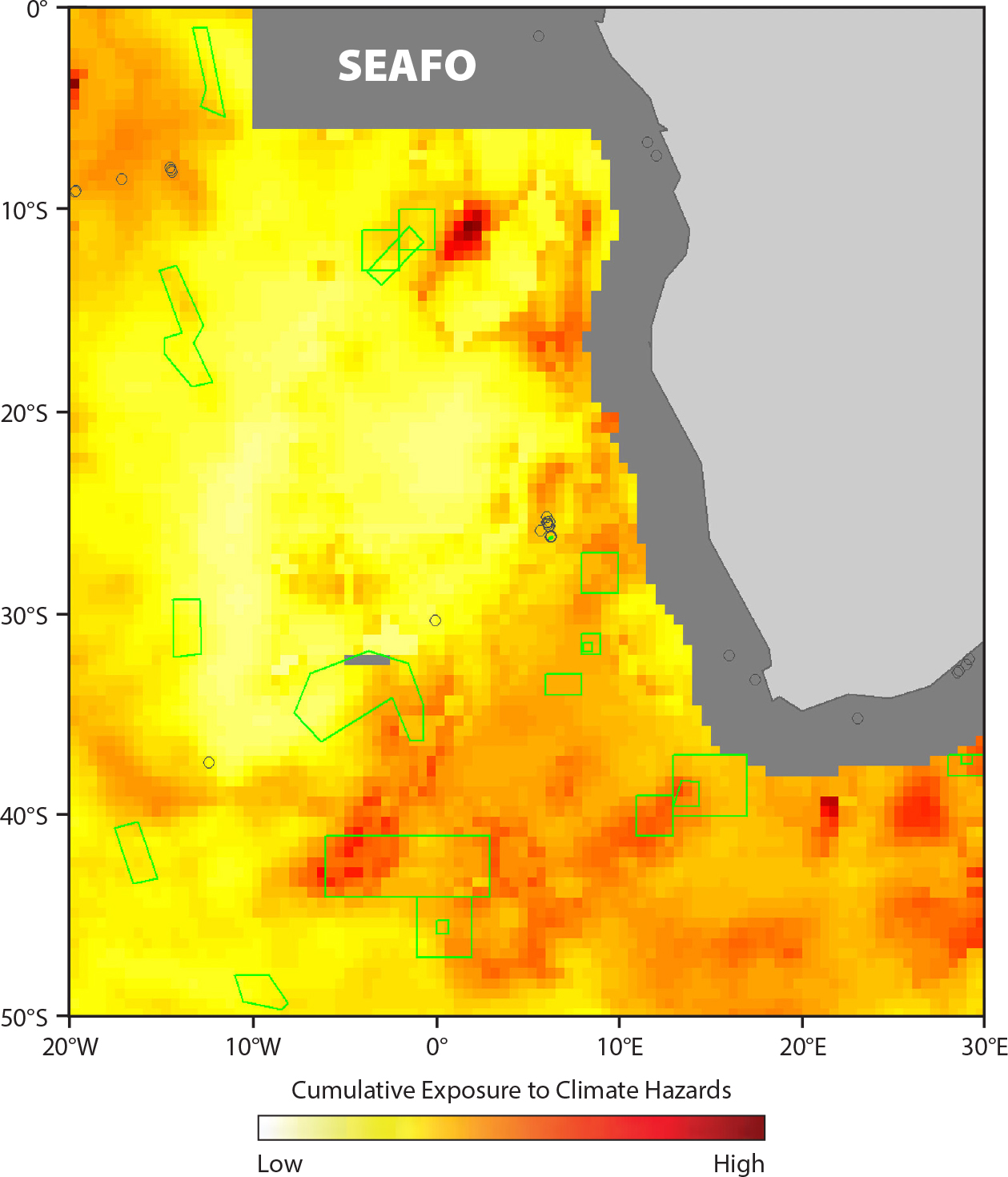
Knowing how to manage fisheries or seabed mining in the face of climate change is made more difficult by uncertainties in the climate models and by not knowing biological climate tolerances, thresholds, and adaptation capacity. Still, across all human economic sectors exploiting the deep ocean, the application of climate Earth system models to project changes in the deep sea (Sweetman et al., 2017) can help to identify not only areas that are important now but those that may become important in the future (Johnson and Kenchington, 2019).
The acquisition of baseline data and the preparation of an EIA are prerequisites for seabed mining, new bottom fishing locations, and potentially other activities in international waters. Because baseline conditions are shifting under climate change, and these could be confused with the impacts of human exploitation of the deep ocean, it is important to build climate-relevant environmental measurements not only into spatial management but also into baseline assessments, and to continue monitoring these parameters as extraction activities occur, in both impacted and protected areas. This will enable identification of cumulative impacts and avoid misinterpretation of direct impacts.
Deep Ocean Stewardship
Meeting the challenge of sustainable blue economies under climate change in the deep sea will require increased understanding by ocean scientists about which observations can help meet regulatory and governance needs, where these observations are most needed, and in what form they can be used by managers and policymakers. Effective stewardship of the deep ocean, balancing conservation with sustainable development, will require accelerated exploration and expanded observing activities, with key roles for international collaboration, capacity building, and possibly philanthropic support. At present, climate science is largely disconnected from applied environmental science and conservation biology. Some of the needed connections can be made by international initiatives, such as the Deep Ocean Observing Strategy (DOOS), which integrates deep-observing activities to address scientific and societal needs; the Deep Ocean Stewardship Initiative (DOSI), which brings together scientific expertise across disciplines and sectors to inform environmental management of the deep ocean; and the International Network for scientific investigation of DEEP-sea ecosystems (INDEEP), which focuses on biological advances.
The challenges of the coming decade will require networks such as these, along with additional deep-sea stakeholders (industry, civil society, conservation interests), to work together through international initiatives such as Sustainable Development Goal 14, Agenda 2030, the UN Decade for Ocean Science for sustainable development, and new policy instruments such as the BBNJ treaty (Levin et al., 2019). But beyond science diplomacy, society must consider if, where, and when sustainability is possible in the deep ocean. Can we develop climate-resilient, deep blue-green economies? To achieve sustainability, we should adopt holistic, climate-conscious environmental policies. Climate can be incorporated into spatial planning, while we reduce the potential for cumulative climate and industry impacts by embarking on aspects of a circular economy, where demand for resources from the deep ocean is reduced by innovation in recycling and reuse, design and manufacturing, efficient distribution, extended product lifespans, lowered consumption, and alternative materials (Thompson et al., 2018). There are many opportunities for scientists to engage with these challenges, from filling the vast array of gaps in research knowledge to synthesizing data and translating findings for policymakers.
Conclusions
The deep ocean offers a repository of genetic potential that ecosystems need in order to adapt to a changing ocean. Conserving this potential may help ensure a healthy ocean for centuries to come. This genetic diversity also offers potential solutions for climate remediation—from CO2 scrubbing enzymes in hydrothermal vent fluids to microbial sequestration of methane. The intersection of climate change and human disturbance is likely to reduce ecosystem resilience, alter times and trajectories for recovery from disturbance, or impose new (non-analog) conditions. Addressing these cumulative effects will involve resolving the disconnect between climate science and societal needs for the deep half of the planet. Science offers insight into where to make new deep-ocean observations and which essential ocean variables should be measured to advance deep-sea sustainability. By integrating climate resilience into ecosystem-based management of the deep ocean, and into the development of new resources, we can contribute to climate goals and understanding as well as to ocean sustainability, and the conservation of biodiversity and ecosystem services (Wüstemann et al., 2017).
|
