
Full Text
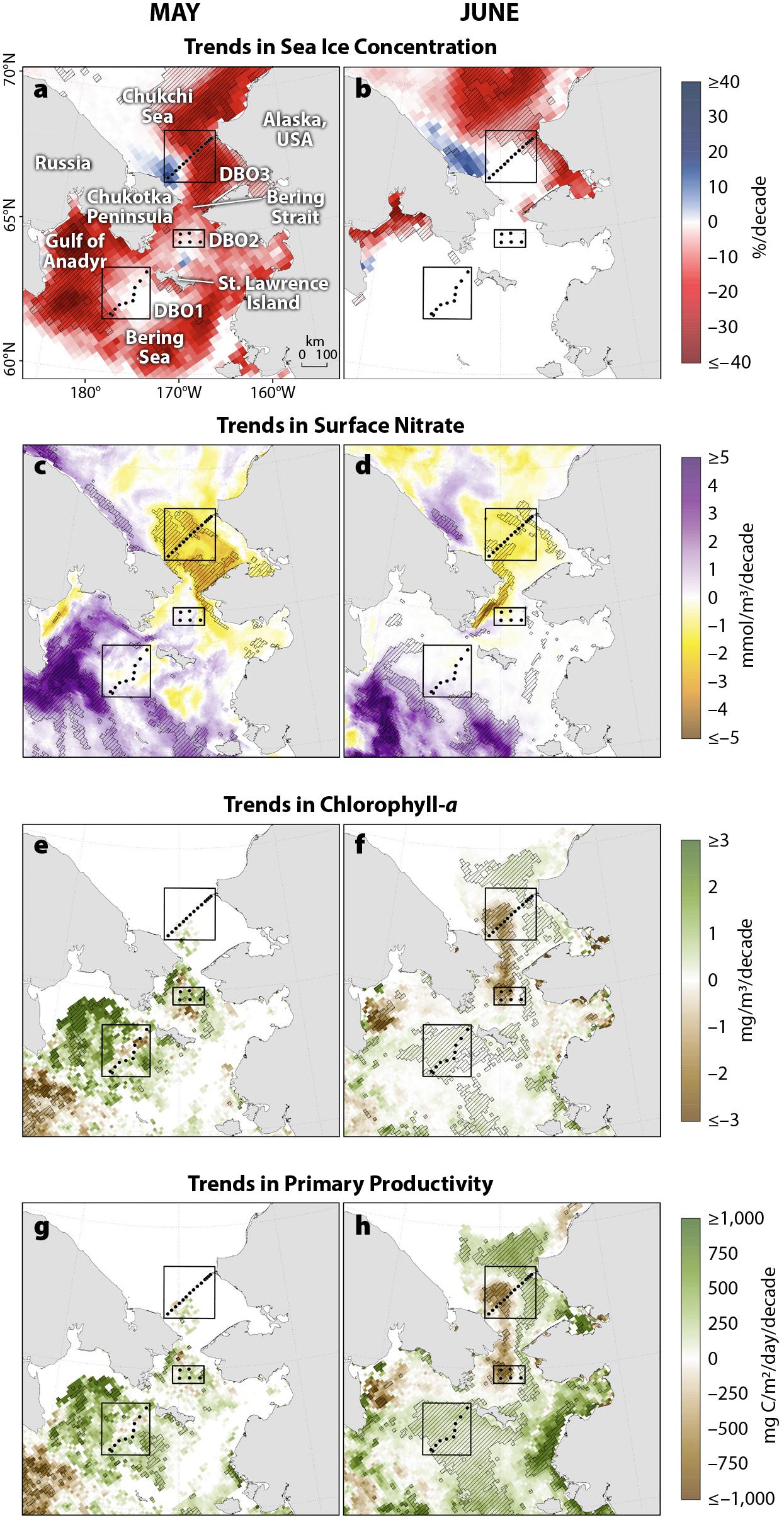
The shallow (~50 m deep), narrow (~85 km wide) Bering Strait is the sole marine link between the Pacific and Arctic Oceans and represents a critical northward throughflow of freshwater, nutrients, and heat into Arctic waters from lower latitudes (Woodgate and Peralta-Ferriz, 2021). Three water masses enter the Chukchi Sea through the Bering Strait from the Pacific: Anadyr Water (AW), Bering Shelf Water (BSW), and Alaskan Coastal Water (ACW) (Coachman et al., 1975). The western Bering Strait in particular has long been known to be a region of consistently high primary productivity throughout the spring and summer open-water season (Sambrotto et al., 1984; Springer and McRoy, 1993; Brown et al., 2011). This productivity is sustained through the delivery of high-nutrient AW waters via the northern branch of the bifurcated Bering Slope Current (Clement Kinney et al., 2009, 2022; Lowry et al., 2015; Pickart et al., 2016) that also causes the Chukchi Sea to the north to be one of the most productive shelves in the Arctic (Hill et al., 2018). Western Bering Strait waters are clearly differentiated from lower productivity waters observed in the eastern Bering Strait that are characterized by relatively low-nutrient, freshwater-dominated ACW (Woodgate and Aagaard, 2005; Lee et al., 2007). However, time series of satellite observations over the last two decades have revealed statistically significant early season (June) declining trends in chlorophyll-a concentrations and primary productivity in the western Bering Strait. In particular, June chlorophyll-a concentrations have declined by ~58%, and June primary productivity has declined by ~34% over the 2003–2020 period. These declining trends appear to be associated with reductions in sea ice cover and increases in primary production upstream in the Gulf of Anadyr during May, with potential implications for decreased nutrient availability downstream in the western Bering Strait during June.
To investigate recent biological change in the Bering Strait, we compiled a satellite-based time series of chlorophyll-a concentrations derived from Aqua-Moderate Resolution Imaging Spectroradiometer (Aqua-MODIS) calibrated radiances using two algorithms: the OC3m algorithm that was developed at NASA Goddard Space Flight Center (GSFC) and makes use of band ratios and in situ measurements (O’Reilly et al., 1998) and the CI algorithm that makes use of reflectance differences in conjunction with a model (Hu et al., 2012). The data are made available by the Ocean Biology Processing Group and were downloaded from the GSFC Distributed Active Archive Center (DAAC) at https://oceandata.sci.gsfc.nasa.gov/MODIS-Aqua/Mapped/Daily/4km/chlor_a/. Chlorophyll-a concentration data were also combined with sea surface temperature data and additional data sets to derive net primary productivity using a broadly utilized algorithm (Behrenfeld and Falkowski, 1997) that has previously been employed to report changes across the Arctic region (Frey et al., 2021). Monthly chlorophyll-a and primary productivity data were only utilized where sea ice concentrations were <10% and were otherwise reported as missing data. For further context, we investigated sea ice concentration data obtained from the Special Sensor Microwave/Imager (SSM/I) and Special Sensor Microwave Imager/Sounder (SSMIS) passive microwave instruments, calculated using the Goddard Bootstrap (SB2) algorithm (Comiso et al., 2017a,b). Modeled surface nitrate concentrations were obtained from the Regional Arctic System Model (RASM; e.g., Clement Kinney et al., 2020). For all data sets (chlorophyll-a, primary productivity, sea ice, and surface nitrate), monthly time series were compiled for May and June, and the Theil-Sen median decadal trends for each month (2003–2020) were calculated, with statistically significant (p<0.1) trends identified using the non-parametric Mann-Kendall test for monotonic trend (Mann, 1945; Kendall, 1975). The Theil-Sen median trend uses a robust non-parametric trend operator that is particularly well suited for assessing the rate of change in noisy and/or short time series (Hoaglin et al., 2000), which in this study is 18 years. For those data sets that include missing data (chlorophyll-a and primary productivity), we show only those trends for pixels that had at least 71% of the time series present (or in the case of this study, 13 of the 18 time steps). This requirement ensures that only robust trends are reported, given that the “breakdown bound” for the Theil-Sen trend is 29% (meaning that unknown or potentially “wild” values would have to persist for more than 29% of a time series in order to affect the overall trend values; Hoaglin et al., 2000).
Increasing trends in marine primary productivity across the Arctic owing to shifts in sea ice cover, seawater temperatures, and nutrient availability have been widely reported (Arrigo et al., 2008; Pabi et al., 2008; Arrigo and van Dijken, 2015; Clement Kinney et al., 2020; Lewis et al., 2020; Frey et al., 2021). In contrast to those reports of large-scale increases in primary productivity, Figure 1 identifies an important and unusual regional location of early season (June) declines in productivity, with potential implications for nutrient and carbon delivery downstream (northward) across the Chukchi Sea shelf. During May (over the 2003–2020 period), we observe strong declines in sea ice concentration in the Gulf of Anadyr (Figure 1a) with increases in surface nitrate concentrations (Figure 1c), and these are in turn associated with increasing trends in both chlorophyll-a (Figure 1e) and primary productivity (Figure 1g). However, during June, we observe (and model with RASM, not shown) strikingly strong and spatially cohesive declines in chlorophyll-a (Figure 1f) and primary productivity (Figure 1h) downstream of the Gulf of Anadyr in the western Bering Strait along the coast of the Chukotka Peninsula. By June, sea ice has typically already exhibited seasonal breakup in the Bering Strait region (Frey et al., 2015), and we see no trends in sea ice cover in the western Bering Strait (Figure 1b). Significant declining trends in June surface nitrate concentrations (Figure 1d) geographically mirror the observed declines in chlorophyll-a (Figure 1f) and primary productivity (Figure 1h). It is important to note that the potential for increased presence of subsurface chlorophyll maxima (as a result of deepening nutriclines) may be challenging to quantify seasonally via satellite data in the Chukchi Sea (Arrigo et al., 2011; Ardyna et al., 2013; Brown et al., 2015). Nonetheless, we hypothesize that because of the May declines of sea ice in the Gulf of Anadyr and resulting increases in May chlorophyll-a/primary production in that region, available nutrients downstream in the western Bering Strait during June are depleted, and chlorophyll-a/primary productivity therefore have declined over time there as well. In particular, in the western Bering Strait (within the region designated as statistically significant for June chlorophyll-a concentrations; Figure 1f), June chlorophyll-a concentrations have changed by approximately –58% (from 4.2 mg/m3 to 1.8 mg/m3), and June primary productivity has changed by approximately –34% (from 2,418 mg C/m2/day to 1,606 mg C/m2/day). These shifts represent chlorophyll-a trends of –1.52 mg/m3/decade and primary productivity trends of –477.8 mg C/m2/day/decade. However, increases in chlorophyll-a and primary productivity in the western Bering Strait primarily during September (not shown) counteract these June decreases, so overall annual primary productivity rates in this region are not significant. Thus, while annual productivity may not have changed substantially, observed shifts in the seasonal distribution of productivity may indeed have profound consequences for marine ecosystem functioning across this region.
|
|
Despite measurements of overall, large-scale increases in primary productivity across the Arctic Ocean over recent decades, heterogeneity in shifts of nutrient availability to upper ocean waters across the region has also led to a spatial mosaic of both increases and decreases in productivity (Juranek, 2022, in this issue). For example, while earlier sea ice retreat can result in stronger blooms in Arctic shelf regions, increased sea ice melt can also result in reduced production in portions of the central Arctic owing to enhanced stratification (Song et al., 2021). Furthermore, moored sensor-based measurements of dissolved inorganic nitrogen (DIN) in bottom waters in the northern Bering Sea indicate high interannual variability but an overall decline of ~50% over the 2005–2017 period, with strong correlations of late summer/early fall DIN resulting in primary productivity downstream on the northern Chukchi shelf the following May (Mordy et al., 2020). Likewise, the early season declines in primary productivity in the western Bering Strait found in this study should undoubtedly have important consequences for the further downstream delivery of carbon and otherwise excess nutrients to the Herald Canyon and western/central Chukchi Shelf regions, important hotspots for biological productivity in the Arctic (Arrigo et al., 2012, 2014; Linders et al., 2017; Li et al., 2019). Changes in the seasonal and spatial distribution of spring phytoplankton blooms in the Pacific Arctic will also likely have important effects on pelagic-benthic coupling in a region with historically high benthic biomass and large populations of seabirds and marine mammals that depend upon benthic prey for survival (Grebmeier et al., 2006, 2018).
The observations of change in the western Bering Strait reported here provide an important example of the heterogeneity of ecosystem responses to climate change, where primary productivity does not always increase with declines in sea ice cover. Moreover, it is important to consider how environmental changes such as sea ice decline can have vital impacts on ecosystem functioning not only locally but also through resulting impacts on nutrient delivery downstream along a conveyor belt system of ocean currents.