Introduction
As a climate-sensitive region where surface air temperature is warming at a pace more than double that of the rest of the globe (Taylor et al., 2017, Jacobs et al., 2021), the Arctic is undergoing profound change. Extensive loss of sea ice area in all months of the year (Stroeve and Notz, 2018) is affecting regional albedo and radiative heat budgets. Decreased sea ice extent and persistence is also allowing increased exchange between the upper ocean and the atmosphere, with enhanced transfer of momentum from atmospheric cyclones and storms, and greater exchange of heat in areas previously covered by ice (Crawford and Serreze, 2017; Serreze et al., 2009; Screen et al., 2011). Thinning ice and increased extent and duration of seasonal open water is increasing phytoplankton primary productivity (PP) in previously light-limited regions (e.g., Arrigo and van Dijken, 2015). Meanwhile, enhanced stratification from increased upper ocean freshwater content in deep basins restricts nutrient replenishment from deep waters, limiting PP and the exchange of gases between the atmosphere and surface waters (McLaughlin and Carmack, 2010; Haine et al., 2015; Carmack et al., 2016).
These shifting baselines in the physical system are already driving changes in the biogeochemical cycling of nutrients and carbon throughout the Arctic Ocean in both predictable and less predictable (or even counterintuitive) ways (e.g., Bates and Mathis, 2009; Tremblay et al., 2015). Bathymetry, stratification, seasonal vs. perennial ice coverage, exposure to storms, degree of river and terrestrial influence, and effects of advection from adjacent regions all determine regional-scale responses. Consequently, Arctic Ocean subregions (i.e., shelves vs. deep basins and areas positioned at Arctic gateways vs. those situated at interior locations on circulation pathways) are forced by a unique mélange of drivers, and their responses vary in both sign and magnitude. Hence, while the long-term decrease of sea ice from the Arctic Ocean is a unifying trend, the ocean’s biogeochemical responses are not singular, but rather a suite of complex, regional-scale trajectories. Here, key aspects of biogeochemical change are highlighted through the lens of a foundational currency, nitrogen, which functions as the primary limiting nutrient controlling PP in the Arctic Ocean and thus is at the heart of many of the biogeochemical changes occurring throughout the region. Insights that emerge from an Arctic Ocean-wide budget of nitrogen, as well as those gleaned from understanding the regional-scale dynamics underlying integrated, Arctic Ocean net change, are discussed as are the consequences of altered nutrient dynamics for the air-sea exchange of CO2 in the Arctic Ocean.
Changing Nutrient Supply in the Arctic Ocean
Water column nutrient distributions comprise a fundamental control on photosynthesis and, hence, the PP that forms the foundation of Arctic Ocean ecosystems. PP, and more specifically, net community production (the fraction of PP that is not respired by heterotrophs in surface waters) also facilitates the sequestration of CO2 in the Arctic Ocean as the carbon contained in organic matter settles to a depth where, upon subsequent oxidation, the resulting CO2 generated is separated from the atmosphere. The major limiting nutrient controlling primary production in the Arctic Ocean is nitrogen, as inorganic nitrogen (hereafter referred to as dissolved inorganic nitrogen, DIN, which includes the sum of nitrate, nitrite, and ammonium species) is typically found with phosphorus in a molar ratio much lower than the canonical Redfield stoichiometry of 16:1 (Codispoti et al., 2013; Tremblay et al., 2015). The deficiency of DIN in Arctic waters can be understood in the context of Arctic Ocean circulation and connectivity to other basins: low N:P waters from the subarctic Pacific Ocean advect into the Arctic Ocean (Yamamoto-Kawai et al., 2006; Tremblay et al., 2015), while additional DIN losses occur within the Arctic Ocean by denitrification on Arctic shelves (Figure 1; Chang and Devol, 2009). Surface waters in much of the ice-free Arctic Ocean exhibit depleted surface DIN inventories quickly after ice retreat, leading to nutrient limitation of PP and oligotrophic status during the rest of the open water season when light is abundant (Figure 2). Climate-related changes to the availability of DIN in surface waters therefore have particular significance for primary productivity and biological pump functioning.
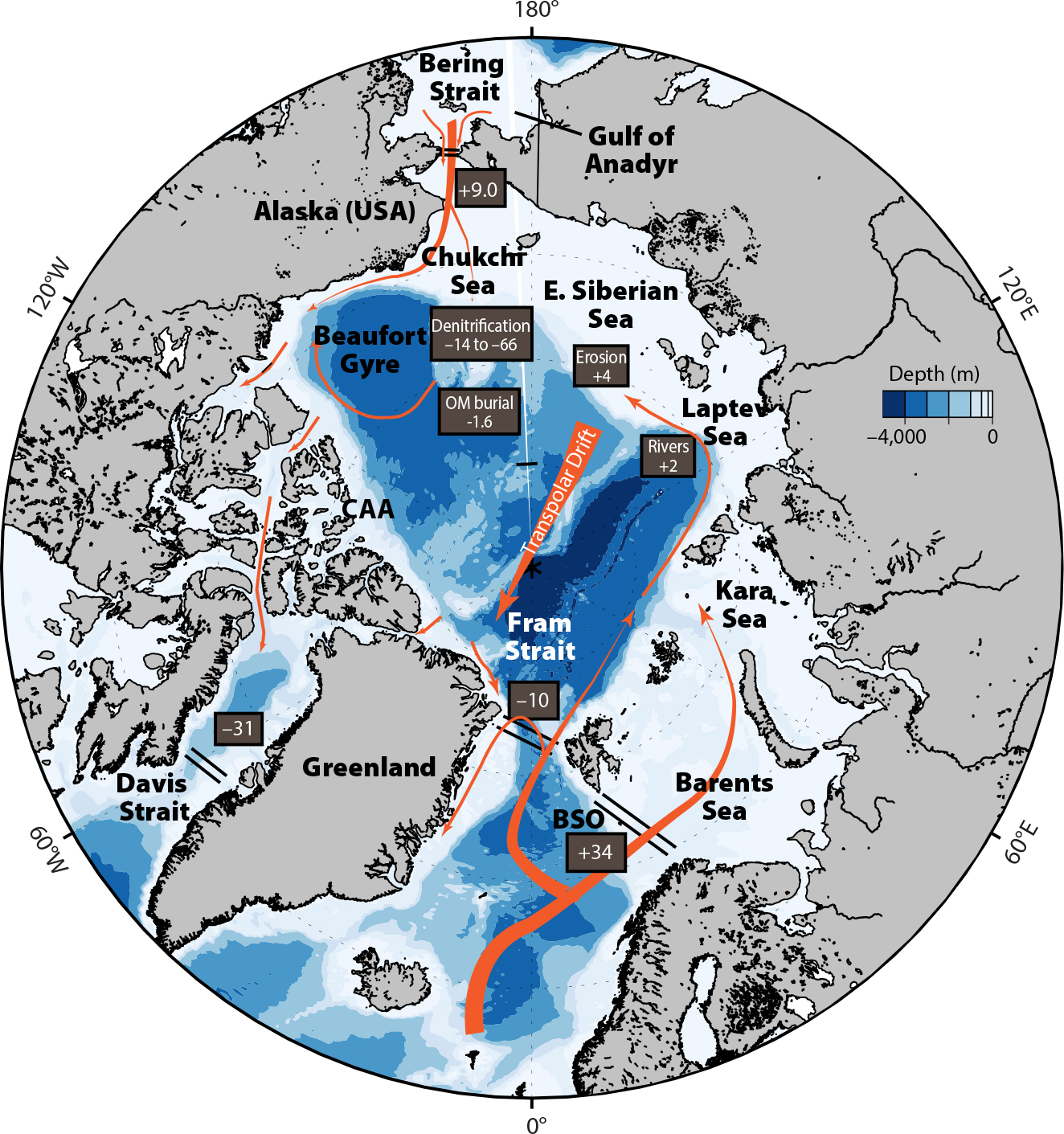
FIGURE 1. Map of the Arctic Ocean showing its gateways at Bering Strait, Fram Strait, Davis Strait, and the Barents Sea Opening (BSO). Major regional seas characterized by inflow shelves (Chukchi and Barents Seas), interior shelves (Beaufort, Kara, Laptev, and East Siberian Seas), and outflow shelves (Canadian Arctic Archipelago, CAA) are also indicated. Orange arrows provide a basic conceptual representation of major circulation pathways referred to in the text. Depth-integrated net dissolved inorganic nitrogen (DIN) transport fluxes through Arctic Ocean gateways are shown in boxes (positive values indicate net flux into the Arctic Ocean, negative values indicate flux out) as reported in Torres-Valdés et al. (2013). Estimates of internal DIN sources and sinks as discussed in the text are also indicated, with positive/negative values indicating DIN sources/sinks, respectively. All quantities are indicated in kmol N s–1. > High res figure
|
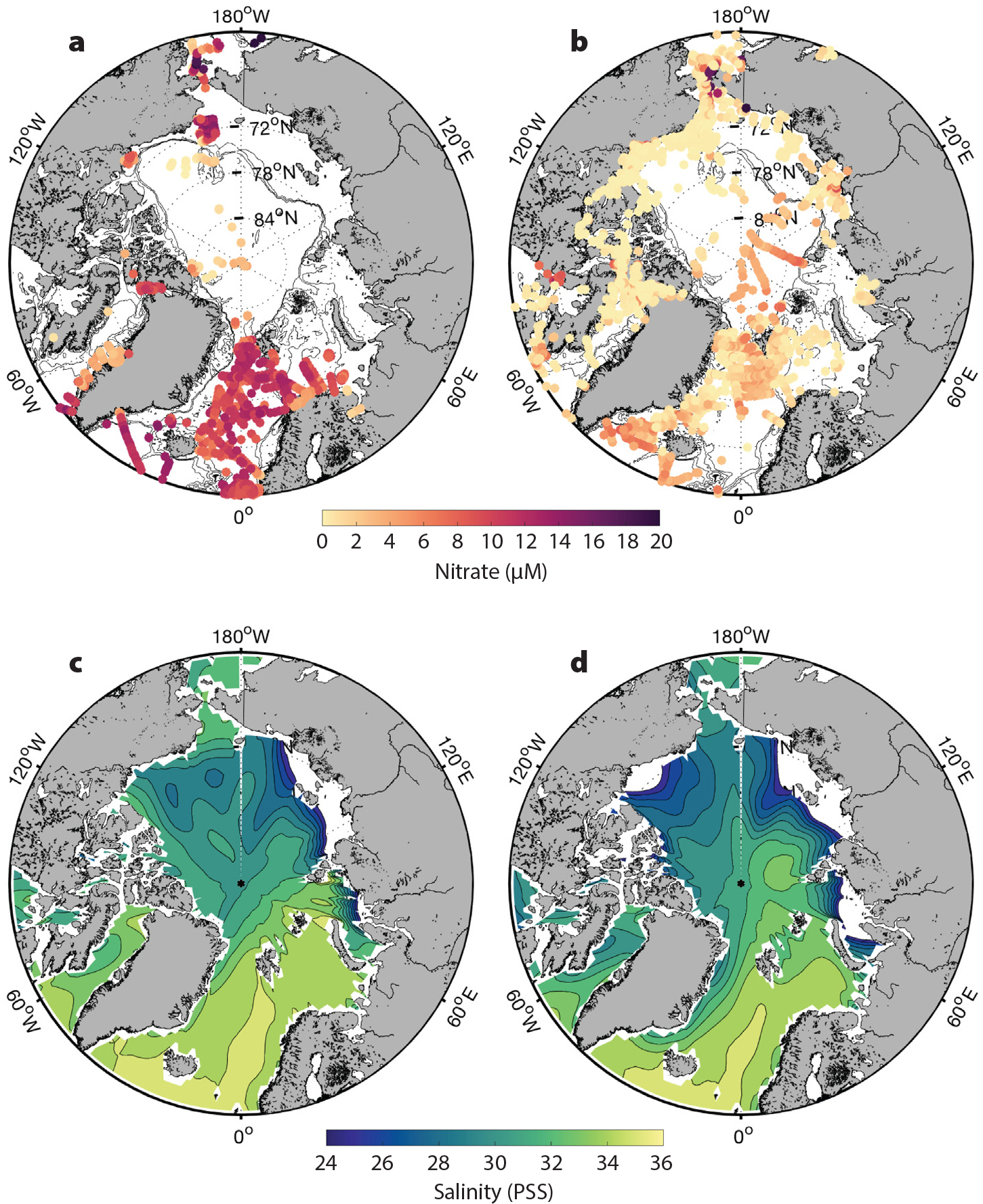
FIGURE 2. Seasonal patterns of nutrients and salinity in the Arctic Ocean. Nutrient data were compiled from Codispoti et al. (2013) as well as additional sources reported in Randelhoff et al. (2020). Salinity is from a seasonal climatology based on the World Ocean Database (Boyer et al., 2014). (a) Winter (November–March) surface (<10 m) nitrate concentration. (b) Post-bloom (August–September) surface nitrate concentration. (c) Winter (January–March) salinity climatology. (d) Summer (July–September) salinity climatology. The nutrient scale has been attenuated to make regional differences more discernible. > High res figure
|
Recent pan-Arctic remote sensing studies indicate an approximate 60% increase in Arctic PP between 1998 and 2018 in open water areas and recognize that greater availability of light and nutrients is fueling this increase (Ardyna and Arrigo, 2020; Lewis et al., 2020). Declining ice cover, increasing open water area, and decreasing sea ice persistence all clearly contribute to more light availability for photosynthesis (e.g., Pabi et al., 2008). Nutrient concentration and flux data are generally not available at sufficient spatial and temporal resolutions to quantify the importance of various mechanisms that increase nutrient supply. Consequently, hypotheses concerning PP increases fueled by changing DIN supply are largely based on inference. As an example, remote sensing indicates that some of the largest increases in PP and chlorophyll concentration occur in the Barents and Chukchi Seas (Lewis et al., 2020), regions known as Arctic “inflow” shelves (Carmack et al., 2006). These regions are situated at Arctic gateways where prevailing circulation advects water masses (and the nutrients they contain) from adjacent subarctic regions (Figure 1). Recent increases in water transport through Arctic gateways (Årthun et al., 2012; Woodgate, 2018; Polyakov et al., 2020) have been hypothesized to increase DIN supply and fuel observed PP and chlorophyll increases (Lewis et al., 2020).
In addition to these remote nutrient inputs from adjacent subarctic seas, a suite of local delivery mechanisms spurred by a changing physical environment are also likely to impact nutrient availability in sunlit waters (Tremblay et al., 2015). These mechanisms are governed by regionally specific physical characteristics (bathymetry, relative ice cover, stratification, wind patterns, and degree of terrestrial influence), as well as factors related to differences in the biological community (grazing rates, community composition); thus, they are spatially variable and operate on a spectrum of inherent timescales. Together, these factors influence both the degree to which nutrients are seasonally replenished in winter and the degree to which episodic nutrient fluxes occur during the ice-free season (Carmack and Chapman, 2003; Pickart et al., 2013; Randelhoff and Sundfjord, 2018; Randelhoff et al., 2020). Mobilization of terrestrial and shelf-derived material from increased river discharge, thawing permafrost, and enhanced coastal erosion also plays an important role in certain regions (Frey and McClelland, 2009; Le Fouest et al., 2013; Terhaar et al., 2021).
A Baseline DIN Budget for the Arctic Ocean
To provide important context for how DIN supply and availability may be changing in various subregions of the Arctic Ocean, it is helpful to first start with a zoomed out, pan-Arctic scale view of how known DIN sources and sinks contribute to this ocean’s baseline budget. The budget approach, a tried-and-true tool in the biogeochemistry playbook, identifies important knowledge gaps and helps provide important context regarding potential sensitivities to perturbations. Multiple attempts to construct DIN budgets for the Arctic Ocean have been undertaken in the last 50 years, often with spatially and temporally sparse data (see Torres-Valdés et al., 2013, and references therein). The most recent and comprehensive effort to date used a model of depth-resolved circulation and measured nutrient profiles from Arctic gateway regions in summer to constrain the DIN inputs and outputs via transport (Torres-Valdés et al., 2013). This analysis found that the major net sources of DIN to Arctic waters were via inflow gateways at the Bering Strait (9.0 kmol s–1) and the Barents Sea Opening (34 kmol s–1; Figure 1). The Fram Strait and Davis Strait gateways also had substantial DIN inputs, but they were offset by large nutrient outputs in equatorward currents. Fram Strait was a net exporter of DIN (–10 kmol s–1), with a balance of inputs (53 kmol s–1), primarily via the West Spitsbergen Current, and outputs (–63 kmol s–1) from the East Greenland Current. The net DIN transport in Davis Strait (–31 kmol s–1) was dominated by the outflow on the western side (–38 kmol s–1), with a weak inflow on the shallow waters of the eastern side (6 kmol s–1). Perhaps more importantly, the sum of all DIN inputs and outputs at the gateways was 1 kmol s–1, indistinguishable from zero, given the methodological uncertainties.
A near-zero net DIN transport is an intriguing result. It suggests that the nutrient budget is balanced with respect to transport, which indicates that there cannot be additional, internal DIN losses within the Arctic Ocean without additional sources. Otherwise, DIN inventories would deplete over time. However, there are a few important caveats. The Torres-Valdés et al. (2013) analysis relied solely on summer transport and nutrient profiles (primarily from a single season in 2005, with some sensitivity studies); thus, seasonal and interannual variability in nutrient fluxes was not captured. As discussed in the following section, the advection of nutrients through Arctic gateways in winter months along with interannual variability in DIN transports are likely critical for setting the inventory of nutrients within the Arctic Ocean. To fully close the nutrient budget requires resolving coupled transport and nutrient fluxes over timescales relevant to the ocean’s circulation, observations that are not yet available.
Nonetheless, it is still useful to evaluate the implications of a net zero transport of DIN through the gateways. Internal system losses of DIN are well documented, and they could not be sustained indefinitely in the absence of additional sources if net transport of DIN were negligible. Loss of DIN by microbially mediated denitrification (conversion of fixed and bioavailable DIN to N2 and N2O) is estimated to be a substantial internal sink term of 14 to 66 kmol N s–1 (–6 to –29 Tg N yr–1; Chang and Devol, 2009; Figure 1). Denitrification is particularly prevalent on shallow Arctic shelves that receive a high flux of organic matter (Chang and Devol, 2009; Granger et al., 2018). Additional loss of DIN is expected through sedimentary burial of organic matter. The majority of organic matter produced via PP is respired back to DIN in surface or subsurface waters, but a small fraction escapes oxidation and is buried (primarily on shallow Arctic shelves and adjacent continental slopes, where the settling time is reduced); estimated burial is 0.7 Tg N yr–1 (3.7 Tg C yr–1; Stein and Macdonald, 2004), which equates to 1.5 kmol N s–1. These internal sinks are partially offset by additional DIN and dissolved organic nitrogen (DON) from terrestrial sources, that is, mobilized by rivers and coastal erosion within the Arctic. These sources affect regional biogeochemical cycling but are believed to be small at the scale of the Arctic Ocean: ~1.5–1.7 kmol N s–1 (Le Fouest et al., 2013; Torres-Valdés et al., 2013). DON flux through Arctic gateways may also represent an important source, but at present it is poorly constrained (Torres-Valdés et al., 2013; Tremblay et al., 2015).
The knowledge gaps that emerge from the large-scale Arctic Ocean nutrient budget point toward areas where there are clear research needs. Resolving nutrient inputs at Arctic gateways over a full annual cycle, along with quantifying interannual variability or trends, is of utmost importance for understanding how PP and biologically mediated CO2 uptake may change in the Arctic Ocean. Better constraint of DON transport/utilization and reduction in the uncertainty of the denitrification sink might also help to bring the budget toward closure. However, the budget analysis also provides important context for understanding what is known about changing sources and sinks, as well as coupled physical and biogeochemical processes that do not act as sources or sinks on an Arctic-wide scale but do impact regional DIN availability in the upper layers where PP and CO2 uptake occurs. The next few sections tackle what is known regarding changing nutrient supply via Arctic Ocean gateways, changing nutrient supply via rivers, and changing upper ocean nutrient availability from physical processes operating over a range of spatial and temporal scales.
Changing Nutrient Supply at Arctic Inflow Shelves
Arctic Ocean gateway nutrient fluxes calculated by Torres-Valdés et al. (2013) relied on data from summer 2005, but observations from Arctic inflow shelves suggest that transport is changing significantly in these regions. At the Bering Strait gateway, a ~50% increase in the volume transport has been observed from the 1990s through 2014 (i.e., 0.7 Sv to 1.1 Sv; Woodgate, 2018), leading some to hypothesize that this corresponds to increased DIN input to the areas immediately downstream (e.g., Ardyna and Arrigo, 2020; Lewis et al., 2020). However, the DIN flux (mass/time) is a product of both the volume transport (volume/time) and DIN content of various water masses (mass/volume) entering Bering Strait. Higher-nutrient water is derived from outer slope waters of the Bering Sea, and in particular waters that circulate in the Gulf of Anadyr to the southwest of Bering Strait (see sidebar by Frey et al., 2022, in this issue). Terrestrial-origin fresh waters conveyed north by the Alaska Coastal Current on the eastern side of Bering Strait tend to be low in nutrients (Codispoti et al., 2013). Long-term trends in salinity monitored at Bering Strait indicate that the transport has freshened significantly, particularly in winter (Woodgate and Peralta-Ferriz, 2021), which suggests that a direct correlation between transport and DIN flux cannot be presumed.
Several lines of evidence suggest that delivery of DIN through Bering Strait may in fact have been decreasing in the past decade. Moored sensor-based observations of near-bottom nitrate concentrations in the northern Bering Sea in late summer/early fall indicate an overall 50% decline over the 2005–2017 period (from ~20 µM to ~10 µM), with a rebound in 2018–2019 (Mordy et al., 2020). These late summer/early fall nitrate concentrations were also found to be highly correlated with those on the northern Chukchi shelf in mid-May, which roughly corresponds to the timing of sea ice retreat and associated spring phytoplankton bloom in this region. In a separate analysis reported in this special issue, Frey et al. (2022) found a decline in remotely sensed PP in western Bering Strait waters typically influenced by high-nutrient Anadyr water. Anomalously high PP in May in the Gulf of Anadyr, hypothesized to be driven by earlier ice retreat, mirrored a decrease in PP in downstream waters of the western Bering Strait in June (34% over 2004–2010), suggesting that nutrients were being consumed in the northern Bering and depleting nutrients that would normally allow PP to occur downstream in the Chukchi Sea (Frey et al., 2022, in this issue).
On the other side of the Arctic Ocean in the Atlantic gateway region, the temporal trend in DIN flux is similarly unclear. Observations suggest a doubling of warm, Atlantic-origin water in the Barents Sea—a phenomenon termed the “Atlantification” of the European Arctic sector (Årthun et al., 2012). The heat content of this water mass has been implicated in the northward migration of the seasonal ice zone (Oziel et al., 2017); in addition, as a major source of nutrients to the region, this water mass might be presumed to support higher rates of PP on the Barents Sea inflow shelf (Henley et al., 2020). However, the degree to which the nutrient content of Atlantic water can be utilized by phytoplankton is influenced by stratification and ice cover, which interact with local wind forcing to set seasonal nutrient replenishment in winter as well as intermittent pulses of nutrients into the system during the open water season (Figure 2; Slagstad et al., 2015; Wiedmann et al., 2017; Henley et al., 2020). The depth of mixing controls the inventory of DIN available for PP in the stratified surface layer; thus, even though Atlantic water may convey a reservoir of nutrients into the Arctic Ocean, it has little influence on Arctic biogeochemical cycling unless it reaches surface waters. An ocean biogeochemical model run under a future warming scenario suggests a decrease in productivity in the southern Barents Sea inflow region over the next century due to enhanced thermal stratification, which reduces nutrient replenishment in winter (Slagstad et al., 2015). In addition, a decline in the nitrate concentration of inflowing Atlantic water in the Barents Sea has been observed over the 1990–2010 period; variations in the source region of waters feeding into the Barents Sea (due to climate-ocean responses to the North Atlantic Oscillation) may play a role in this trend (Rey, 2012; Oziel et al., 2017).
In addition to nutrient-based controls on PP on inflow shelves, the importance of other processes that regulate biomass, including advection of phytoplankton and grazers from adjacent regions (Vernet et al., 2019; Wassmann et al., 2019), should be considered. Recent biogeochemical modeling in the Barents Sea and Fram Strait regions suggests that advection of phytoplankton from south to north along major currents supports a substantial proportion of biomass and resulting PP (Vernet et al., 2019). The importance of advected vs. in situ production is seasonally and spatially variable, but the upshot is that this advection allows more northerly regions to maintain much higher rates of PP than they would with no advective inputs. This advected biomass ultimately amounts to a 0.76 Tg C yr–1 supplement of organic carbon (and its stoichiometric equivalent of organic N) to the Arctic Ocean north of Svalbard, with more southerly subregions within the Atlantic water inflow receiving higher subsidies (Vernet et al., 2019). Advection of grazers (e.g., copepods and microzooplankton) northward also regulates existing biomass and is an important control on PP in inflow shelf regions (Lavrentyev et al., 2019; Wassmann et al., 2019). Thus, physical and ecological factors that influence grazer communities (surface warming, changes in advective transport, changing spatial patterns of phytoplankton biomass) will ultimately influence Arctic Ocean PP trends as well.
Increasing Influence of Terrestrial Nutrient Sources
Recent studies also implicate nutrients supplied by coastal erosion and rivers as having an increasingly important role in supporting observed PP increases and influencing coastal biogeochemical cycling in the Arctic Ocean. Increased river discharge, thawing permafrost with deepening active layers, and enhanced shoreline erosion due to a loss of buttressing ice in fall and winter all intensify the land-ocean exchange of material (Frey and McClelland, 2009; McClelland et al., 2012). One recent modeling analysis that sought to quantify the impact of terrestrial sources on the Arctic Ocean indicates that DIN supplied by coastal erosion and rivers (estimated as 1.6 Tg N yr–1 and 1.0 Tg N yr–1, respectively) supports one-third of Arctic PP on an annual basis (9%–11% for rivers and 19%–41% for coastal erosion; Terhaar et al., 2021). Consistent with prior work (e.g., Peterson et al., 2002; Frey and McClelland, 2009; Holmes et al., 2012), the majority of the terrestrially derived DIN sources were focused in the Eurasian Arctic (East Siberian, Kara, and Laptev shelves), where the Yenisey, Lena, and Ob’, the three largest Arctic rivers by annual discharge and the fifth, sixth, and thirteenth largest rivers globally, respectively, are located. However, the N supply was only estimated to support a biomass increase (new production) of 17 Tg C yr–1, while the simulated increase in productivity was eight times the biomass increase (140 Tg C yr–1). Hence, the large modeled PP response was mostly from continued recycling of the initial (modest) DIN input (i.e., regenerated production; Dugdale and Goering, 1967). This finding is consistent with prior analyses that also found modest contributions of river-derived nutrients to new production (Tank et al., 2012; Le Fouest et al., 2013) due to the carbon-rich and nitrogen-poor nature of Arctic rivers (Holmes et al., 2012). The distinction between new vs. regenerated PP is important because these two types of PP influence carbon and nutrient cycling differently. Regenerated production is a zero-sum process. It does not involve a net biological uptake of CO2 because regeneration and reuse of N contained in organic matter also regenerates CO2. New production is not zero sum, and can support biomass transfer to higher trophic levels, thus fueling Arctic ecosystems. Alternatively, organic matter produced during new production can sink to subsurface water masses that are out of reach of seasonal mixing horizons; therefore, the carbon and nitrogen contained in settling organic matter becomes isolated from further biogeochemical cycling in the surface environment.
Observations from the recent Arctic GEOTRACES mission provide additional evidence of an increasing signature of terrestrial and shelf inputs into Arctic Ocean waters. Kipp et al. (2018) found that an increase in radiotracer activities (228Ra and 226Ra) in the Transpolar Drift of the central Arctic Ocean (Figure 1) in 2015 relative to 2007 is indicative of increased shelf-based inputs from East Siberian and Laptev Sea shelves). They hypothesized that disturbance of a large 228Ra reservoir in shelf sediments by enhanced wind-driven mixing over a longer open water season was the primary driver of these changes. These findings are broadly consistent with those of a coastal erosion modeling study (Terhaar et al., 2021) that found the largest modeled increases in marine PP from terrestrial and sedimentary nutrient sources occur on the East Siberian, Laptev, and Kara shelves.
The Kipp et al. (2018) study as well as aforementioned river and coastal erosion studies all paint a picture of Eurasian shelves that are “interior” in Arctic Ocean circulation pathways (Figure 1; Carmack et al., 2006) and heavily influenced by large Arctic rivers as sites of intensified land-ocean biogeochemical cycling and sediment mobilization. This unique regional character is important to note because, as described above, these regions exhibit a different suite of responses to Arctic Ocean change than are seen in other regions.
Changes to Seasonal Nutrient Replenishment
In addition to nutrient sources from rivers and transport through gateways, processes that affect the depth distribution of nutrients over an annual cycle are also critical influences on biogeochemical nutrient and carbon cycling in the Arctic Ocean. In seasonally and perennially ice-free waters, pre-bloom nutrient inventories at the surface are well correlated with patterns of annual PP throughout the Arctic Ocean (Figure 2; Tremblay and Gagnon, 2009; Tremblay et al., 2015; Randelhoff et al., 2020). The association is somewhat intuitive because DIN inventory amassed during winter mixing determines the reservoir available to phytoplankton when sea ice seasonally thins and retreats and light levels become sufficient for growth (Codispoti et al., 2013; Tremblay et al., 2015; Randelhoff and Sundfjord, 2018). However, the degree of winter replenishment varies across Arctic shelves and deep basins, governed by regional stratification and the depths of winter mixing, and the nitracline (Wassmann and Reigstad, 2011; Tremblay et al., 2015; Randelhoff et al., 2020). Typically, shallow shelves nearest to nutrient sources at the gateways exhibit more robust replenishment while open waters overlying deeper bathymetry and strong stratification show weakest replenishment (see winter nitrate concentrations in Figure 2). For example, the Chukchi Sea in the Pacific Arctic sector is characterized by a broad, shallow shelf (ca. 50 m) and a relatively shallow nitracline (ca. 20–40 m); in winter, storms and convective mixing completely erode the seasonal stratification, and the nutrients accumulated in the near-bottom layer from remineralization of organic matter over the summer are redistributed throughout the water column (Figure 3; Pacini et al., 2019; Mordy et al., 2020). The Barents Sea shelf is deeper (ca. 200 m), but this region has also historically been characterized by extensive replenishment of DIN by convective and storm-driven mixing in fall/winter (Figure 2; Slagstad et al., 2015; Randelhoff et al., 2020). The strong replenishment contributes to the very high rates of PP in these locales during ice retreat in spring (Hill and Cota, 2005; Codispoti et al., 2013; Matrai et al., 2013).
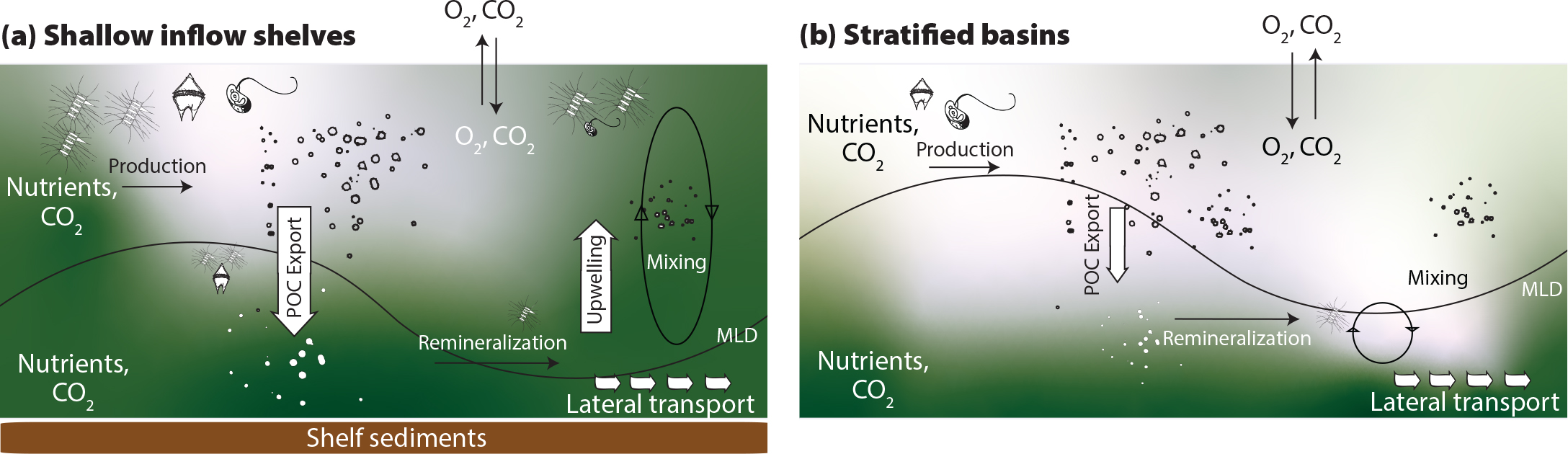
FIGURE 3. Conceptual diagram of nutrient replenishment mechanisms on (a) shallow inflow shelves, and (b) stratified deep basins. Green shading indicates relative nutrient availability. On inflow shelves, advective nutrient inputs from lower latitudes and strong replenishment due to storm-induced and winter mixing supports high rates of primary production (PP) and organic matter export. Stratified interior shelves and basins have weaker winter nutrient replenishment and storm-induced mixing, with consequent lower PP rates. MLD = mixed layer depth. > High res figure
|
In contrast, the depth of the nitracline in the deep Canada Basin of the central Arctic Ocean exceeds that of typical winter mixing depths (McLaughlin and Carmack, 2010; Carmack et al., 2016; Randelhoff et al., 2020). In regions of the high-latitude central Arctic Ocean that have transitioned from perennial to seasonal ice cover, nutrient replenishment is also weak, but a lack of complete DIN drawdown during the vegetative season in these regions indicates that light limitation still plays an important role in limiting PP (Figure 2; Randelhoff et al., 2020); hence, at present, the role of seasonal nutrient replenishment is not as important here, but this pattern is expected to change with continued declines in ice cover (Slagstad et al., 2015).
The degree of seasonal nutrient replenishment is certainly sensitive to future climate forcing. For example, biogeochemical modeling indicates that PP in the southern Barents Sea will be restricted by future warming and enhanced thermal stratification (Slagstad et al., 2015). Observations suggest that enhanced stratification in the Beaufort gyre has already depressed the nitracline, limiting the resupply of DIN to surface waters (McLaughlin and Carmack, 2010), consistent with both the low PP rates typically observed in this region and an observed shift to smaller picoplankton that are better adapted for nutrient-limited conditions (Li et al., 2009). Meanwhile, warming surface temperatures may fuel enhanced microbial loop activity and facilitate less export of organic material and more regeneration of nutrients in the surface layer (Kirchman et al., 2009). Biogeochemical modeling also predicts further decreases in PP in the Beaufort gyre in the future (Slagstad et al., 2015). The fundamentally different conditions on inflow shelves and in deep basins with respect to declining ice cover and seasonal nutrient replenishment typify the mosaic of Arctic Ocean subregion responses to warming-induced physical system change. On inflow shelves, decrease in ice coverage over winter months facilitates mixing and momentum transfer, whereas in deep basins enhanced haline stratification limits seasonal replenishment. Again, these differences demonstrate that understanding the unique character of Arctic Ocean subregions is critical to understanding biogeochemical responses to climate-driven changes.
Episodic Nutrient Delivery by Storms and Wind Events
While changes in transport and winter replenishment set total nutrient inventories available to phytoplankton in the upper water column for early season growth, episodic pulses of nutrients contributed by regional wind-forcing and/or current-bathymetry interactions can be important for maintaining productivity throughout the post-bloom, summer, open-water season when nutrients are scarce (Pickart et al., 2013; Ardyna et al., 2014; Nishino et al., 2015; Wiedmann et al., 2017). Because these mechanisms help to relieve nutrient limitation and support continued lower trophic level production over a lengthening growing season, they may be increasingly important in a warming Arctic.
Enhancement of shelf break upwelling by the expansion of the seasonal melt zone has long been recognized as an important mode by which PP might be enhanced in a warming Arctic (Carmack and Chapman, 2003), though at more spatially and temporally reduced scales. Reduced or minimal ice cover at continental shelf breaks facilitates the transfer of wind momentum; when prevailing high and low atmospheric pressure centers result in directional, upwelling-favorable winds and shelf break depths are shallow enough to constrict flow and allow horizontal divergence (Randelhoff and Sundfjord, 2018), nutrient-rich waters are brought to the surface. This is particularly true along the Beaufort Sea shelf break and in the vicinity of Barrow Canyon in the Northeast Chukchi Sea (Pickart et al., 2013). While much of the Eurasian Arctic sector is characterized by relatively deep shelves, conditions for upwelling may be favorable at the comparatively shallow Laptev Sea shelf break (Randelhoff and Sundfjord, 2018).
Analysis of long-term wind and mooring data in the vicinity of the Beaufort shelf break indicates that upwelling can be induced by moderate easterly winds (threshold of 6 m s–1) and that the frequency of upwelling-favorable events has likely increased in recent decades (Pickart et al., 2013). The estimated upward DIN flux associated with these events could support significant rates of new production (average of ~400 mmol C m–2 per storm) if all supplied DIN is converted to biomass, but the extent to which this occurs is presently unknown. Remotely sensed and ship-based observations do indicate clear PP response to coastal upwelling events (Pickart et al., 2013), and the long-term satellite chlorophyll record notably indicates increased concentrations at both the Beaufort and Laptev Sea shelf breaks (Lewis et al., 2020).
The duration of upwelling events likely plays a role in their overall impact—longer events will allow more time for phytoplankton communities to respond and draw down available DIN inventories. Retentive circulation features facilitated by current-bathymetry interactions (e.g., Okkonen et al., 2011) may extend PP responses beyond the lifetime of the initial wind forcing. Recent work also suggests that shear and instabilities related to frequent changes in wind forcing can induce higher rates of cross-isopycnal nutrient flux in the Chukchi Sea (Beaird et al., 2020). In contrast to the “reversible” nutrient fluxes facilitated by temporary upwelling events, these “non-reversible” turbulent nutrient fluxes enhance transfer of N from nutrient-rich bottom waters to shallower mid-water column depths where light is sufficient to fuel photosynthesis.
More generally, there is growing recognition that turbulent and storm-induced nutrient fluxes away from shelf breaks may also play an increasingly important role in supporting higher PP in the Arctic Ocean as the seasonal ice zone and open water growing seasons expand. An analysis of satellite chlorophyll from 1998 to 2012 found increased prevalence of fall blooms throughout the Arctic Ocean attributed to greater frequency of high-wind events during open water conditions in September and October (Ardyna et al., 2014). The most significant increases in fall bloom occurrence were on inflow shelves (Chukchi, Barents), Eurasian interior shelves (Siberian, Laptev, Kara), and ice-free portions of the central Arctic. More recently, a biogeochemical modeling study found that high-frequency winds can facilitate higher Arctic Ocean primary productivity by two main nutrient-delivery mechanisms: first, and most significant, was enhanced and earlier deepening of mixed layer and nutrient entrainment in fall when light was still sufficient to allow phytoplankton blooms to occur; second, a prolonged mixing period in winter enhanced nutrient inventories that could fuel spring productivity (Castro de la Guardia et al., 2019). Thus, an increase in mixing associated with high wind events as the open water season expands is likely to manifest in both spring and fall. Evidence of earlier spring blooms has also been noted in some areas, including the Canadian Arctic Archipelago, Baffin Bay, and the Kara Sea (Kahru et al., 2011).
Implications for Regional Changes in Air-Sea CO2 Exchange
The coupling of carbon, nitrogen, and phosphorus in biologically mediated processes (Redfield et al., 1963) means that changing patterns of nutrient supply and consequent changes to marine PP are inextricably linked to carbon cycling as well. However, changes to the physical system (warming, increased river discharge and increasing freshwater content in the Arctic Ocean basins, sea ice loss, changing wind speeds) also exert strong control on carbon exchange at the atmosphere-ocean and the terrestrial-ocean boundaries. While a full review of the Arctic Ocean carbon cycle and important changes is not in the purview of this review (readers are directed to some excellent reviews on the topic, e.g., Bates and Mathis, 2009, and Olsen et al., 2015), here, the focus is on the ways that inorganic carbon cycling, and in particular exchanges of CO2 between atmosphere and ocean, are responding to the physical and biological system changes mentioned in previous sections.
Changes to Arctic Ocean CO2 Uptake Due to Changing Primary Productivity
Regions of the Arctic Ocean that have seen marked increases in PP over the last several decades, in particular the Arctic gateway inflow shelves, will also act as strong biologically mediated sinks for CO2 on a seasonal basis as organic matter is produced in the surface and exported to depth. As exported organic matter is respired at depth, seasonal stratification prevents mixing of accumulated respiratory CO2 to the surface where it would otherwise outgas, much as it restricts the resupply of DIN that also accumulates (Figure 3). Input of CO2 associated with respiration of exported organic matter at depth also contributes to seasonal undersaturation of calcium carbonate minerals (i.e., corrosivity; Bates and Mathis, 2009). Historically, this biologically mediated organic carbon pump has helped the Chukchi and Barents Seas to maintain strongly undersaturated CO2 at the surface and has facilitated these areas functioning as regions of enhanced ocean uptake of CO2 from the atmosphere, while interior shelves and deep basins with low PP rates represent much weaker sinks (Bates and Mathis, 2009; Yasunaka et al., 2016; Pipko et al., 2017). The recently reported increases in PP on some Arctic shelves therefore have the potential to enhance oceanic uptake of CO2 so long as they are associated with export of material to depth and not with regenerated production, where CO2 is alternately consumed and released by photosynthesis and respiration, respectively (Dugdale and Goering, 1967; Tremblay et al., 2015). For example, the substantial proportion of PP fueled by riverine and coastal erosion sources of DIN found by Terhaar et al. (2021) would only modestly contribute to enhanced biological CO2 uptake because the majority of the PP in that study was determined to be regenerated. In contrast, a combination of modeling and observations suggests an increase in continental shelf PP and biological CO2 uptake over a longer growing season on inflow shelves (Tu et al., 2021) and indicates the importance of high rates of PP on the Chukchi shelf for offsetting reduction in CO2 uptake associated with surface temperature increases (Ouyang et al., 2020). In Arctic Ocean deep basin surface waters, a reduction of an already weak biologically mediated CO2 sink due to enhanced stratification and associated deepening of the nutricline (McLaughlin and Carmack, 2010) has already been noted (Cai et al., 2010; Else et al., 2013; Ouyang et al., 2020).
PP associated with the previously described shelf break upwelling and fall phytoplankton blooms warrants additional discussion with respect to new/regenerated production and biologically mediated air-sea CO2 exchange. The supply of DIN from deep waters that fuels these blooms also brings excess CO2, leading to significant outgassing, as has been noted in several studies (Mathis et al., 2012; Hauri et al., 2013; Evans et al., 2015). Consumption of DIN at the surface during the phytoplankton bloom helps to mitigate this outgassing by drawing down surface water CO2; hence, the timescale of DIN removal during a bloom helps to set the net source/sink status of CO2 flux during these upwelling events. Whether or not these events represent new vs. regenerated production depends on the depth from which nutrient-rich waters are sourced: DIN supplied from shallow horizons where organic matter produced earlier in the season has been respired would not, in an annual budget sense, be considered new production, while DIN supplied from previously untapped reservoirs would.
Additional potential shifts in phytoplankton community composition in response to warming and other physical system changes may also reduce the efficiency of biologically mediated CO2 uptake in the Arctic Ocean. Already, the Atlantification of the southern Barents Sea and the northward migration of the polar front have been implicated in the proliferation and northward expansion of coccolithophore blooms in this region (Neukermans et al., 2018, and references therein; Figure 4). Formation of CaCO3 shells reduces alkalinity of surface waters and increases the partial pressure of CO2 in surface waters (pCO2 w, where subscript w denotes water), weakening the capacity for air-sea CO2 uptake. Coccolithophores tend to proliferate under conditions expected in this region in the future (i.e., warm waters containing low to moderate nutrients, and shallow mixed layers; Balch, 2018). While coccolithophore blooms are not common in the Chukchi Sea, they have been noted in the adjacent Bering Sea (Napp and Hunt, 2001; Ladd et al., 2018; Figure 4). The overall consequences of these ecological shifts for net regional CO2 uptake will require continued monitoring and ecosystem modeling to resolve.

FIGURE 4. Remotely sensed visible images from MODIS Aqua show coccolithophore blooms in the (a) Barents Sea and (b) Bering Sea. (c) Laptev Sea turbidity seen here is associated with terrestrial material delivered by the Lena delta. (a) and (b) NASA images courtesy of Jeff Schmaltz, MODIS Rapid Response Team, NASA GSFC; (c) NASA image by Norman Kuring/NASA’s Ocean Color Web, using MODIS data from NASA EOSDIS/LANCE and GIBS/Worldview. > High res figure
|
Changes in Abiotic Drivers of Arctic Ocean CO2 Uptake
Warming and freshening of Arctic Ocean surface waters will also impact the source/sink status of some regions, particularly deep basins and some interior shelves where biologically mediated rates of CO2 uptake are low. The expansion of the seasonal ice zone and reduction in sea ice extent in theory presents an opportunity for enhanced ocean uptake of CO2, as cold high-latitude surface waters can now more readily communicate with the atmosphere (Olsen et al., 2015), but warming and freshening of Arctic basins counter this potential. Increasing pCO2 w in the Beaufort Sea over the last few decades, an expected outcome of warming, has already been noted in a number of studies (Cai et al., 2010; Else et al., 2013; Ouyang et al., 2020), resulting in reduced regional CO2 uptake. Seasonal freshwater input from sea ice melt or river discharge (Carmack et al., 2016) additionally reduces the capacity of these waters to buffer against additions/removals of CO2 (by biological or abiotic processes), leading to reduced uptake capacity with additions of CO2 (Rysgaard et al., 2011; Takahashi et al., 2014; Ouyang et al., 2020). Increasing thermal stratification in the Barents Sea is also expected to reduce the CO2 sink in this region in the future. Indeed, observations already indicate increasing pCO2 w and decreasing CO2 uptake in the southern Barents Sea (Yasunaka et al., 2016). This contrasts with the other inflow shelf, the Chukchi, where biotic factors appear to be dominating over warming-related reductions (Ouyang et al., 2020; Tu et al., 2021).
Intensified Land-Ocean Carbon Exchange on Eurasian/Siberian Shelves
Generally, discharges of Arctic Ocean rivers are organic carbon rich and DIN poor (Holmes et al., 2012; McClelland et al., 2012). Thus, while the effect of PP supported by riverine nutrients may be slight, the impact of the organic carbon supplied by rivers on ocean-atmosphere CO2 fluxes can be quite prominent, particularly on interior shelves in the Eurasian Arctic sector where a significant fraction of Arctic Ocean riverine discharge takes place (Anderson et al., 2009; Frey and McClelland, 2009; Pipko et al., 2017). Remineralization of allochthonous organic matter increases pCO2 w, while turbidity associated with increased particle load lowers light penetration and dampens primary productivity (Figure 4; Carmack et al., 2006). These processes are reflected in a gradient toward increasing pCO2 w from west to east across the Eurasian shelves, with low pCO2 w in the Barents Sea driven by cooling of Atlantic water and high productivity rates and increasingly high pCO2 w toward the eastern Siberian Seas where stratification-induced warming of surface waters, high pCO2 w in river discharge, and high rates of terrestrial organic matter remineralization dominate (Anderson et al., 2009; Pipko et al., 2017). The East Siberian Seas still represent a sink for CO2 but are prone to periods of outgassing; it might be expected that with increased mobilization of permafrost and continued warming, these areas might become a more reliable source of CO2 in the future (Anderson et al., 2009). More generally, enhanced stratification from increased surface temperatures and increased river discharge may reduce CO2 uptake capacity on Eurasian interior shelves, although this stratification increase may be countered by increased storm-induced-mixing (Pipko et al., 2017).
Another important trend in this region is a significant increase in alkalinity exported to the Arctic Ocean from the Yenisei and Ob’ Rivers; between 1974 and 2015, alkalinity export by these two rivers more than doubled (from 225 to 642 Geq yr–1 and from 201 to 470 Geq yr–1 for the Yenisei and Ob’, respectively; Drake et al., 2018). Proposed drivers of this increase include increased temperature, deepening of the permafrost active layer, and longer contact time with unweathered mineral surfaces. If similar increases in alkalinity export apply for the two other large Eurasian rivers (Lena and Kolyma), the increase in Arctic Ocean alkalinity has the potential to enhance CO2 sequestration by 3.4 Tg yr–1 (120 Tg C over 1974–2015; Drake et al., 2018). This increase is of the same magnitude as the compiled estimates of regional CO2 uptake in the Kara, Laptev, and East Siberian Seas, which range from 1 Tg C yr–1 to 6 Tg C yr–1 (Bates and Mathis, 2009); hence, increased buffering capacity will be an important determinant of future CO2 uptake in this region.
Conclusions
Changes in the biogeochemical cycling of nutrients through Arctic Ocean sub-regions are inevitable as aspects of the physical system change. DIN availability might increase due to advected inputs at inflow regions, increase due to reduced ice conditions coupled with enhanced storm activity (shelf break upwelling or storm-induced mixing), or decrease as source waters change or stratification limits seasonal replenishment. Hence, there is no single, unified trajectory for Arctic Ocean biogeochemical change—rather, profound regional differences shape a mosaic of trends and outcomes. Changes to nutrient availability already seem to be driving changes in Arctic Ocean PP (Lewis et al., 2020; Terhaar et al., 2021), but what is less clear is the extent to which these trends are driven by new or regenerated production. This new/regenerated distinction has important implications for understanding future changes in Arctic ecosystems, trophic transfers, pelagic-benthic coupling, and capacity for biologically mediated air-sea CO2 exchange.
Given the variety of individual regional responses, observations and modeling are both critical needs for tracking the shifting baselines of Arctic Ocean biogeochemical change, the mechanisms driving them, and implications for the future. Observations must be collected at appropriate spatiotemporal scales to resolve processes of interest; however, this is challenging given the importance of event-driven features (storms) and the need for measurements outside of the easily accessible open water period (to assess seasonal nutrient replenishment trends). Models will also need to adequately resolve mixing processes and their responses in shallow coastal environments and deep basins alike. Finally, given the interwoven functioning of Arctic Ocean physical, biogeochemical, and ecological systems, our ability to understand and predict future change hinges on interdisciplinary coordination from measurement to synthesis.
Acknowledgments
The author is grateful for the tireless efforts of the Arctic research community to understand the multiple facets of Arctic Ocean change and to provide valuable data resources that spur further synthesis and research. The author also thanks the efforts of two reviewers, whose comments greatly improved the manuscript. LWJ received partial support from National Science Foundation awards 1949593 and 1928684 to write this manuscript.

