Introduction
Marine protected areas (MPAs) are designated regions set aside to manage conservation efforts, with the primary aim of preserving and protecting marine life. Effective conservation considers the overall ecosystem functions, encompassing the physical, geological, and geochemical aspects of the habitat, and their relationships with biological communities, as well as the functional relationship among the ecosystems within the MPA and the neighboring undesignated marine areas (e.g., Hays et al., 2020). Preservation efforts also extend to the cultural significance of the marine area and the sustainable use of its resources (Gomez et al., 2021).
Managing an MPA involves balancing multiple—often competing—concerns, such as habitat protection and sustainable use. Effective management must be informed by a strong scientific understanding of an evolving ecosystem, which requires continuous collection of key observations. For MPAs situated in the deep sea, this can be facilitated remotely through sensors delivering time-series observations and recurrent collection of physical samples that help to interpret the continuous sensor data. However, the impacts on the protected area from sensor deployment and data collection as well as of recurring scientific and maintenance expeditions also need to be considered in the MPA management plan (e.g., Cuvelier et al., 2022).
In 1984, the human-occupied vehicle Alvin confirmed the existence of “unusually large” sulfide structures and biological communities supported by hydrothermal venting off the west coast of Canada (Tivey and Delaney, 1986). These structures and communities were localized to the Endeavour Segment of the Juan de Fuca Ridge within Canada’s exclusive economic zone. Upon discovery, and with its fortuitous proximity to coastal ports, the Endeavour Segment became a mecca for scientific research, enabling the dissemination of what some describe as its magical nature and broad recognition in Canadian society of Endeavour’s unique features and their environmental and socioeconomic significance (Tunnicliffe and Thomson, 1999).
Although the size of hydrothermal vent fields is relatively small globally, their ecological significance is high; and even though they are generally located in the remote deep sea, they are threatened by human disturbance (Van Dover, 2012). The process of hydrothermal venting concentrates minerals at the discharge sites, making them ideal candidates for deep-sea mining. The scientific interest they generate can also raise threats of overzealous sampling and other disturbances (Turner et al., 2019).
As a signatory to the Convention on Biological Diversity (1993), Canada resolved to protect 30% of its oceans by 2030. In 2003, Canada began this process by establishing the 97 km2 Endeavour Hydrothermal Vents (EHV) MPA as Canada’s first MPA and the world’s first protected hydrothermal vent site (Figure 1). Established under Canada’s Oceans Act, the primary conservation objectives were to ensure that human activities in the area contributed “to the conservation, protection, and understanding of the natural diversity, productivity, and dynamism of the ecosystem” and that these activities were “managed appropriately such that impacts remained less significant than natural perturbations” (Fisheries and Oceans Canada, 2010).
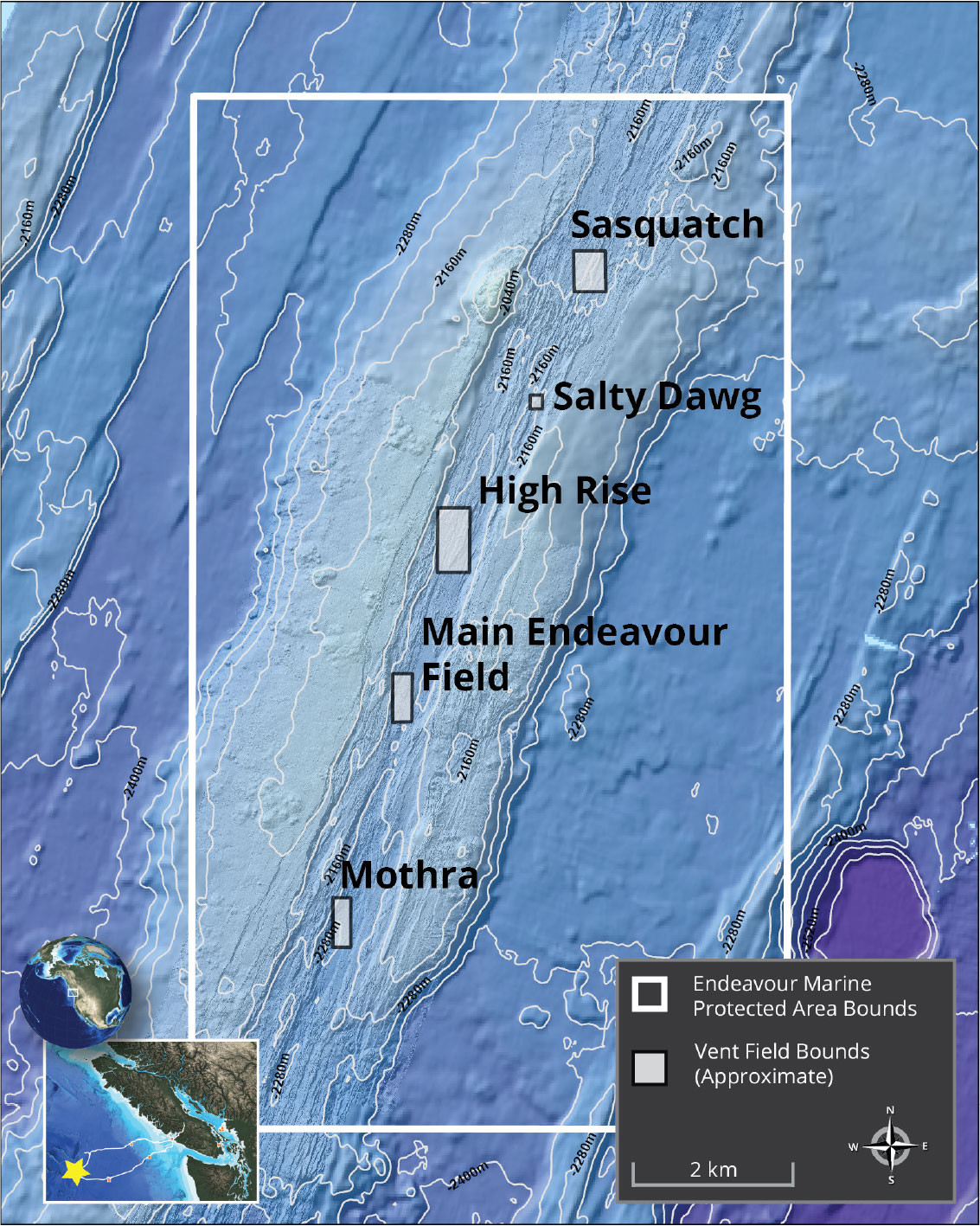
FIGURE 1. The boundaries (white-lined box) and the five main active vent clusters (shaded boxes) of the Endeavour Marine Protected Area are delineated here on a bathymetric map. Coordinate system: WGS 1984 UTM Zone 9N. Image credit: Ocean Networks Canada. > High res figure
|
The EHV MPA is on the Endeavour Segment of the Juan de Fuca Ridge, a section of the global mid-ocean ridge (MOR) system located in the Northeast Pacific Ocean off the west coast of British Columbia. The MOR extends for 70,000 km throughout the global ocean and is where tectonic plates diverge and new oceanic crust is formed. This spreading process results in a permeable seafloor, allowing cold seawater to percolate downward where it is heated by rising magma from the upper mantle. During its subseafloor circulation, the seawater reacts chemically with the surrounding crust and is eventually ejected back into the ocean as mineral-laden, oxygen-depleted, and superheated fluid. The process of mixing with cold, oxygenated seawater leads to a succession of rapid chemical reactions, which form precipitates and creates the chimney-like hydrothermal vents that are the hallmark of the Segment (Figure 2). As the buoyant vent plume rises, the hot metal- and sulfide-rich hydrothermal vent fluid continually reacts with the seawater to create dark, smoke-like, emissions highly enriched in Fe, S, Cu, Ca, and Zn (Feely et al., 1987). The plume rises 200–300 m above the seafloor, at which point it reaches a neutrally buoyant state and spreads with the local oceanic currents as the chemical processes continue (Coogan et al., 2017). This flux of vent fluids plays a major role in maintaining the ocean’s chemical balance. Nearer to the seafloor, chemosynthesis-based biological communities utilize both the energy exchange occurring when these chemical species mix with the oxygenated seawater and the chemical species themselves to form the basis of the hydrothermal vent ecosystems on the seafloor and in the water column (Van Dover, 2000; Burd and Thomson, 2015).

FIGURE 2. This close-up view shows a black smoker chimney at the Main Endeavour Field. Image credit: ONC and CSSF–ROPOS. > High res figure
|
This paper provides an overview of the main geological, biogeochemical, and physical processes at the Endeavour Segment and their roles in regulating the biological communities and habitat structures that host ecosystems at the vents and near the seafloor. We describe highlights of the past 16 years of scientific research and monitoring enabled by the NEPTUNE seafloor cabled observatory that support management decisions for the MPA. Recently, the EHV MPA’s boundaries were repealed and subsumed into the 133,017 km² Tang.ɢ̱wan – ḥačxʷiqak – Tsig̱is (TḥT) MPA. This significantly expanded area is of cultural and economic significance to coastal Indigenous peoples of the west coast of North America and is cooperatively managed by the Council of the Haida Nation, the Nuu-chah-nulth Tribal Council, the Pacheedaht First Nation, and the Quatsino First Nation, together with Fisheries and Oceans Canada (Government of Canada, 2024).
The NEPTUNE Observatory
In addition to its designation as an MPA, the Endeavour Segment was also selected as one of the three Integrated Studies Sites for the US National Science Foundation-funded Ridge 2000 program (Fornari et al., 2012) that attracted significant global scientific attention. Highlighting Endeavour’s scientific value, the proposal for a NEPTUNE cabled observatory was successfully funded, with the primary purposes to understand the spreading, subduction, and faulting of the Juan de Fuca plate, as well as the ecosystems and oceanography off the west coast of Canada. For the purposes of MPA management, the deep-sea observatory enhances observation and monitoring in the area.
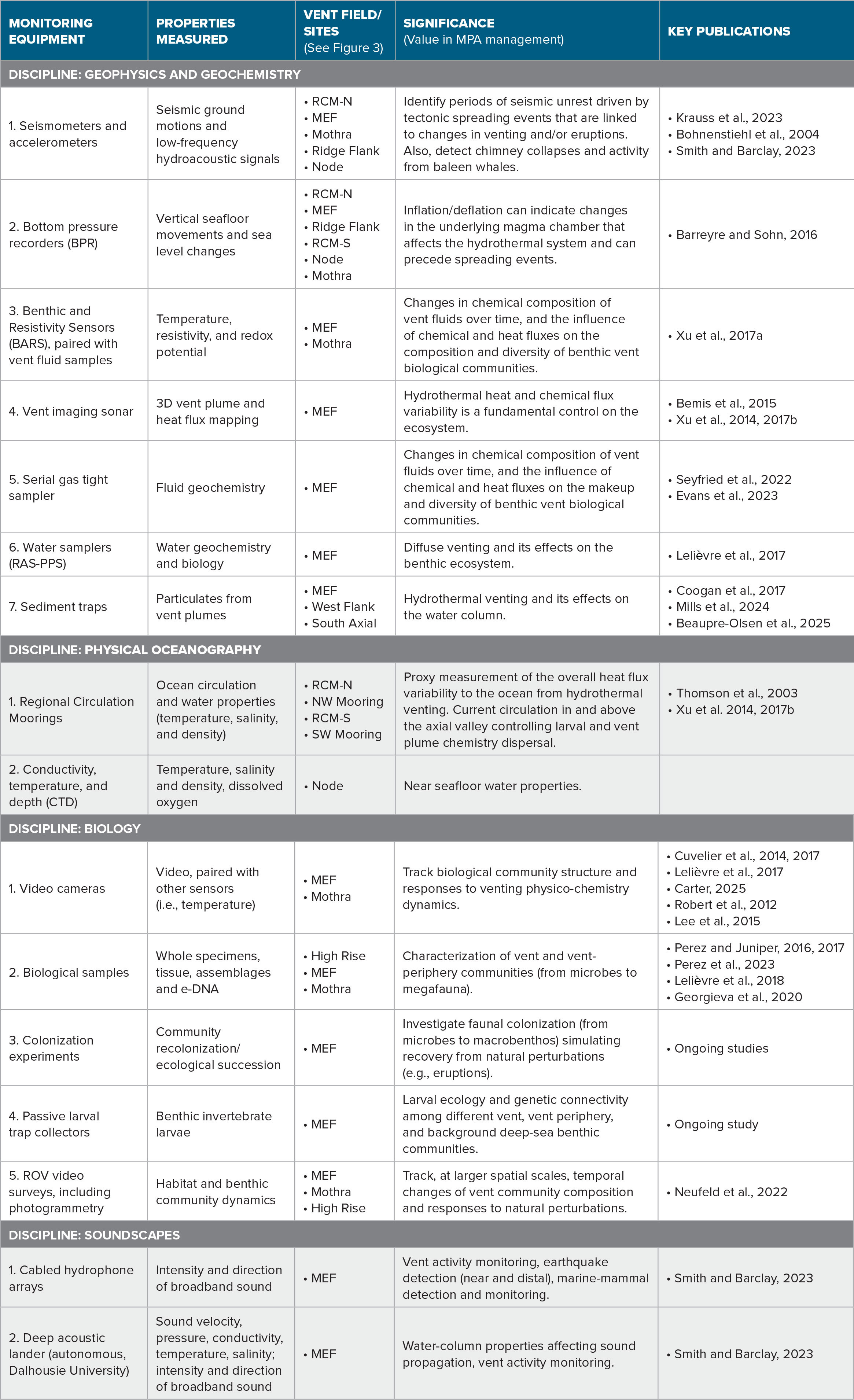
Operated by Ocean Networks Canada (ONC), the NEPTUNE observatory comprises an 840 km fiber-optic cable extending from Vancouver Island across the North American and Juan de Fuca tectonic plates to connect five major node sites to power and the internet (Barnes et al., 2007). The westernmost Endeavour node supports the scientific sensors in the heavily instrumented Endeavour Hydrothermal Vents MPA (Table 1, Figures 3 and 4). Real-time data, archived data, and data products from sensors in the axial valley and on the flanks of the Endeavour Segment of the Juan de Fuca Ridge have been available since 2010 through ONC’s digital infrastructure, Oceans 3.0 (Owens et al., 2022). Regular expeditions using ships and remotely operated vehicles (ROVs) to maintain infrastructure also collect observations and physical samples to ground truth and complement the sensor data. Internet connectivity from the ships allows the Canadian and international science community to participate from shore to conduct experiments and sampling strategies to aid in developing a more complete understanding of the physical, geological, and biological processes of this protected environment (Table 1).
TABLE 1. Summary of major disciplines, sensor technology, and geological, physical, chemical, and biological properties monitored and their scientific and MPA monitoring impacts. MEF = Main Endeavour Field. RCM = Regional Circulation Mooring North and South. > High res table
|
Geology and Seafloor Habitat
The Endeavour MPA hosts hydrothermal vent ecosystems whose formation and persistence are directly linked to the dynamic geological and tectonic processes of the Juan de Fuca Ridge. Understanding this interplay through continuous, multidisciplinary monitoring is foundational to effective MPA management.
The Endeavour Segment is an intermediate-rate spreading center (full rate: ~52 mm yr–1) (DeMets et al., 2010; Krauss et al., 2023) characterized by a 10 km long and 1 km wide axial valley flanked by rift crests rising 100–150 m above the valley floor. Extensive and vigorous hydrothermal venting occurs within the axial valley focused at five main active vent clusters spaced about 2–3 km apart (Figure 1; Kelley et al., 2012). The Main Endeavour (MEF) and Mothra fields have received significant scientific attention and are currently being monitored by sensors connected to the NEPTUNE observatory. The High Rise and Salty Dawg fields (see Figure 1) are designated for minimally intrusive studies and outreach opportunities (Fisheries and Oceans Canada, 2010) and have no cabled sensors.
The uniqueness of hydrothermal venting regions, with respect to other deep-sea benthic habitats, stems from the chemical flux and exchange of heat between the ocean and the seafloor and the geologically rapid change of the seafloor morphology due to local tectonic dynamics. This leads to a chemically and physically extreme environment that hosts the specialized life that has physiologically and biologically adapted to the geologically controlled environment. Although vent ecosystems are rare and their global extent is small, their contribution to the understanding of life and their ecosystem functions and services are significant and are considered ideal candidates for designation as Vulnerable Marine Ecosystems and recommended for Area-Based Management Tools (e.g., Menini and Van Dover, 2019).
The Endeavour Segment features a combination of active and inactive chimneys, edifices, and mounds along its axial valley. The active structures cluster into the five major vent fields with more than 400 inactive structures as well as the diffuse venting sites interspersed among them. Conceptually, they are geologically connected, and the entire ridge segment can be considered a single temporally and spatially varying vent field driven by heat from a continuous axial magma chamber (Jamieson and Gartman, 2020). Over at least the last 2,000 years, there have been no large-scale eruptions with significant lava flows that could bury these vent fields (Jamieson et al., 2013; Clague et al., 2014, 2020). Krauss et al. (2023) attribute this to the degassing of the axial magma chamber, which limits extrusive magmatism and results in the mature chimneys, edifices, and mounds, both active and inactive, that define the Segment. Concurrently, the underlying shallow magma chamber ensures sufficient heat and chemical flux to sustain regular hydrothermal circulation and consistent sulfide structure growth.
Long, continuous time series of physical parameters provide opportunities to reveal complex dynamics and ongoing evolution of the vent system. For example, Barreyre and Sohn (2016) correlated vent fluid temperatures with bottom pressure fluctuations and estimated the permeability of the shallow upflow zones near hydrothermal venting using poroelasticity theory. The study revealed that the Main Endeavour Field (MEF) possesses geospatially distinct shallow upflow zones characterized by different effective permeabilities, which sets it apart from Lucky Strike and the East Pacific Rise, sites with different geological characteristics and spreading rates.
At Smoke and Mirrors, located near the southern Benthic and Resistivity Sensors (BARS; shown in the MEF inset in Figure 3), Barreyre and Sohn (2016) modeled higher effective permeabilities characteristic of a slow spreading center, such as Lucky Strike, with lower heat flux and a thicker extrusive layer that has ample permeable pathways. Just 150 m apart at Grotto (located near the northern BARS shown in the MEF inset in Figure 3), they modeled higher effective permeabilities that are characteristic of a fast spreading center, such as the East Pacific Rise, with higher heat flux and a thinner extrusive layer more frequently paved by volcanic activity.
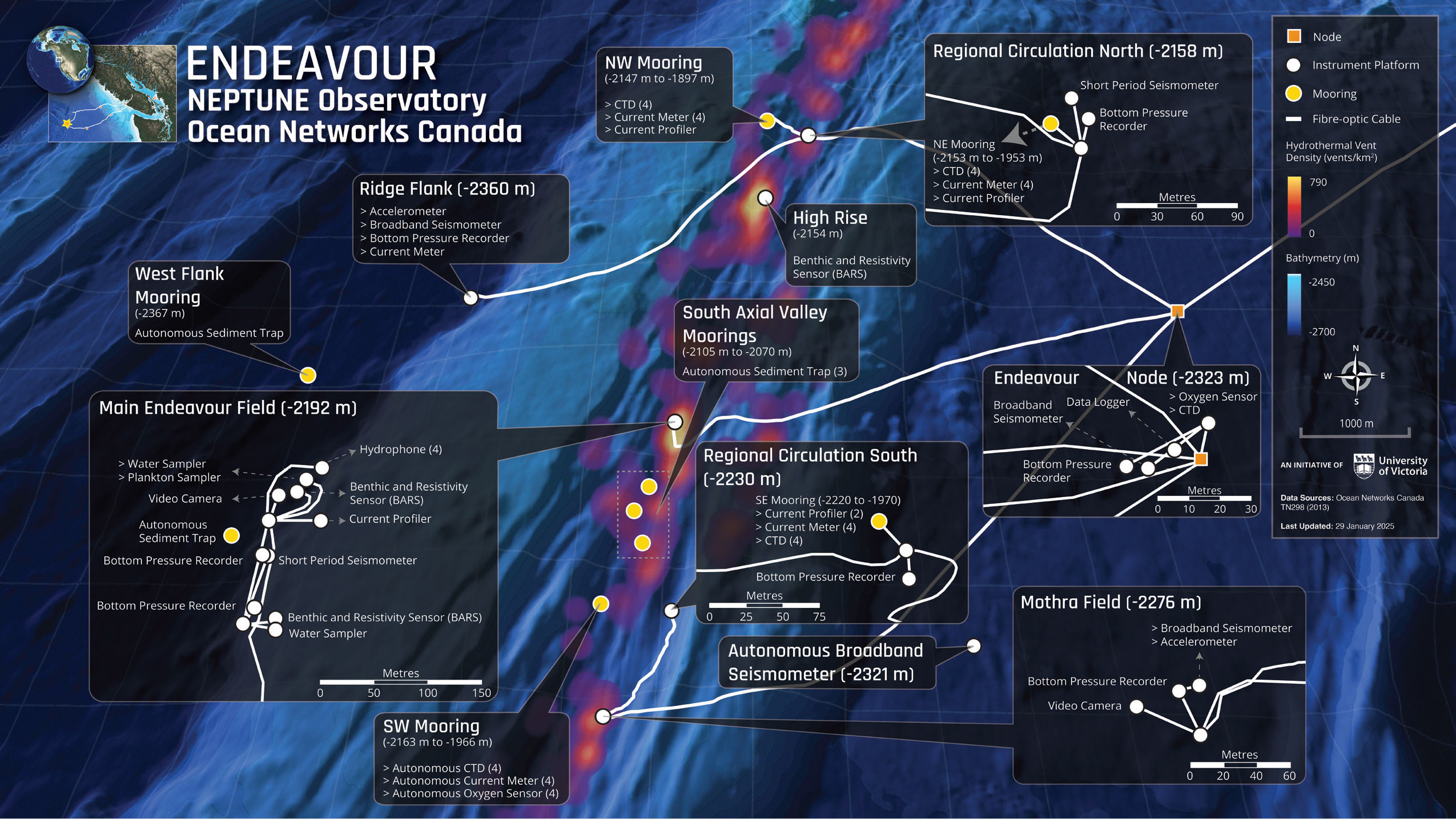
FIGURE 3. ONC infrastructure within the area of the Endeavour vent fields and adjacent ridge flanks. The node (orange square) powers the instrument platforms (white circles) that host scientific sensors connected to the internet via fiber-optic cables (white lines). Moorings—both autonomous and cabled—are shown as yellow circles. Image credit: ONC. > High res figure
|
Rather than attributing these differences to spreading rate, the effective permeability likely varies due to output from the irregular distribution of the underlying magma body. This is corroborated by anecdotal visual evidence from repeated visits showing the southern part of the MEF waning in black smoker output (e.g., Smoke and Mirrors edifice), while the northern part of the field (Grotto edifice) is growing and gaining in vigor. These results and visual observations also imply that the magma supply within intermediate spreading centers can vary in space and time (possibly rapidly) and therefore regionally modify the benthic environment to host biological communities that are more suited to fast or slow spreading centers.
Continuous geophysical monitoring, primarily using cabled seismometers and bottom pressure recorders (BPRs, Table 1), tracks the tectonic activity that can drive these changes in the environment. The real-time data streams permit continuous monitoring of seismic events and seafloor deformation, providing insights into processes influencing environmental stability and enabling timely responses to significant events. Since 2018, heightened seismicity has been observed by Krauss et al. (2023), mirroring precursors to past diking events (1999–2005). This culminated in a notable increase in activity in March 2024, including an M4.1 earthquake and periods with up to 200 events per hour, suggesting the segment may be approaching the next diking event, prompting the scientific community to meet in November 2024 to prepare for a rapid response to a major perturbation of the system.
Understanding the chemical environment driving these ecosystems is critical. Due to the harsh environment of hydrothermal venting regions, geochemical sensors for measuring the continuous temporal variability of the chemistry of fluid emissions are very limited, and scientific research has relied predominantly on laboratory analysis of discrete samples obtained on scientific expeditions. To obtain a continuous time series at high temperature vents, ONC employed a cable-connected BARS to measure temperature, resistivity, and redox potential (eH) of the vent fluids in situ (Table 1). With discrete samples taken at the beginning of the deployment and at the time of recovery, the continuous time series of the sensors’ measurements are used to infer changes in fluid chemistry. However, these sensors reside in black smoker vents with 300°–350°C fluid emission and often do not last a full year between maintenance expeditions. Another method to improve time resolution of the variability of chemical fluxes is to remotely collect discrete samples. Currently, a serial gas tight sampler is deployed alongside a BARS. Its containers can be remotely triggered to collect a time series of 12 vent fluid samples (Seyfried et al., 2022). The timing of the sampling is adapted to changes in seismicity or vent fluid temperatures, allowing correlation between specific geological events and vent fluid chemistry.
The current period of heightened seismicity marks a critical phase for the evolution of the Endeavour Segment. It presents a rare opportunity to observe a potential dike intrusion or spreading event that would offer valuable data for refining models of mid-ocean ridge processes. Studying the Endeavour Segment, with its intermediate spreading rate characteristics, provides a key comparison point between fast- and slow-spreading systems across the globe and other intermediate spreading centers (e.g., the Galápagos Spreading Center). An impending tectonic event may cause significant shifts in hydrothermal output (heat and chemistry), providing a natural experiment to study the resilience and adaptive responses of the specialized vent communities within the MPA. To better capture such an event, Dalhousie University and the University of Washington, in partnership with ONC, enhanced observatory capabilities by deploying five autonomous ocean bottom seismometers in summer 2024; an additional 20 ocean bottom seismometers (including replacements for the 2024 units) are scheduled for deployment in summer 2025. This denser network will improve detection and location of seismicity, providing crucial data for understanding geological and tectonic processes and their impacts on hydrothermal vent ecosystems. It will inform future MPA management strategies regarding natural and anthropogenic disturbances.
Oceanic Environment
Changes in and redistribution of heat and chemical fluxes from the vent fields along Endeavour’s axial valley alter seafloor characteristics, affecting benthic ecosystems as well as the overlying water column and the pelagic ecosystem it hosts. Not only is the seawater chemistry directly altered, but changes in the heat flux from hydrothermal venting affects the local ocean circulation through changes in buoyancy input from the rising hydrothermal plumes. On an axial valley scale, the rising plume generates inflow near the seafloor toward the hydrothermal vents, which facilitates the retention of vent field larvae and plankton. Conversely, the rising plume can also entrain planktonic organisms, moving them up into the water column where along-axis currents can relocate them to vent sites with more retentive circulation, or higher up in the water column where they can be swept away by ambient ocean currents to less hospitable ocean environments (Thomson et al., 2003). If the organisms have the ability to swim vertically or alter their buoyancy, they can use this circulation to move to a preferred location. There is observational evidence of larvae exhibiting this type of behavior at other vents sites (Mullineaux et al., 2013). On the segment scale, the off-axis propagation of the plume alters the chemistry of the ocean (Coogan et al., 2017; Beaupre-Olsen et al., 2025) and has a marked impact on the overlying pelagic ecosystem, enhancing secondary productivity (Burd and Thomson, 2015).
Estimating the flux of hydrothermal fluid and heat along the Endeavour Segment has generally been conducted by observing water property anomalies, either by dense shipborne or autonomous underwater vehicle (AUV) sampling. These observations are inverted to estimate flux using the known temperature of the vent fluid as it leaves the seafloor (Kellogg, 2011), resulting in an overall value of heat flux over the time window of the repeated surveys. With yearly AUV surveys (2004, 2005, 2006), Kellogg and McDuff (2010) identified a transient anomaly over the Salty Dawg vent field, suggesting that there is spatial and temporal variability in hydrothermal flux; however, their temporal resolution made it difficult to determine both the subseafloor causes and the water column effects. As an alternative to annual AUV surveys with their inherent coarse time resolution, four moorings of current meters and water property sensors (CTDs) were installed in the axial valley of the Endeavour Segment. The array is designed to utilize the “sea breeze effect” caused by the rising buoyant plume. This effect relates horizontal currents to the intensity of heat flux from the hydrothermal venting (Thomson et al., 2003); therefore, variability in hydrothermal heat flux is continuously estimated in real time. This four-mooring array also monitors the background bathymetrically modified circulation that controls the mixing and dispersal of the chemical-laden hydrothermal plume (Xu et al., 2013; Coogan et al., 2017; Figure 3), and enables researchers to relate circulation dynamics to ecosystem dynamics (Cuvelier et al., 2014; Lelièvre et al., 2017).
A specialized sonar was developed to image rising plumes in real time after researchers observed that avoidance sonar on submersibles could detect reflections from these plumes (Bemis et al., 2015). The deployment of the Cabled Observatory Vent Imaging System (COVIS; Figure 4) marked a significant technological milestone. COVIS allows for direct tidal-frequency resolution of the total flux from the multiple hot vent orifices that makes up the rising buoyant plume. Utilizing the imagery of the acoustic backscatter off the turbulent fluctuations of the buoyant plume and the Doppler shift of the backscattered signal, researchers were able to estimate the rising plume velocity and the expansion rate and heat flux to the ocean from the hydrothermal venting and its variability through time, and to gain insights into the diffuse low temperature flow (Bemis et al., 2012; Xu et al., 2013, 2014).

FIGURE 4. Artist’s rendering of a selection of ONC’s cabled and autonomous instruments monitoring Endeavour. (a) Regional circulation mooring. (b) Junction box. (c) Bottom pressure recorder. (d) Hydrophone array. (e) Broadband seismometer. (f) Cabled Observatory Vent Imaging Sonar. (g) Passive larval trap collector. (h) Sediment trap. (i) Deep Acoustic Lander. (j) Remotely operated vehicle. (k) Water sampler. (l) TEMPO-Mini ecological module. (m) Benthic and Resistivity Sensors. Image credit: ONC. > High res figure
|
Chemical analysis of hot vent fluid samples collected by a remotely controlled, internet-connected serial gas tight sampler revealed details of the input of nutrient transition metals (e.g., V, Cr, Mn, Fe, Co, Ni, Cu, Zn, Mo) from the oceanic crust to the water column (Evans et al., 2023). These metals play an important role in nutrient-related biological processes. They are essential for the growth of organisms and can be rapidly utilized in near-surface waters and therefore limit growth. Determining the dynamics of chemical flux across the seafloor interface using these types of cabled seafloor samplers informs understanding of the benthic-pelagic coupling that regulates the trophodynamics over regional scales and offers insights into the global role of hydrothermal venting in primary and secondary productivity in the ocean (e.g., Burd and Thomson, 2015; Cathalot et al., 2021).
Soundscape
A significant challenge when monitoring a site like Endeavour is posed by the aggressive environment that can deteriorate instrumentation quickly, especially when placed in the vicinity of the plume. Passive acoustic monitoring (PAM) from hydrophones positioned at a safe distance from the hot and chemically corrosive fluids is used to monitor, in real time, the soundscape at the site for extended periods of time. For example, PAM was applied successfully to the detection and classification of explosive events at volcanically active sites (Chadwick et al., 2008).
Different features of the sounds produced by venting are related to the physical mechanisms producing the sounds. These, in turn, are influenced by physical parameters such as flow rate, chimney height, sound speed, and cavity size (Little et al., 1990; Crone et al., 2006; Smith and Barclay, 2023). Studies aimed at establishing the connection between these parameters and the sounds produced can, in principle, enable the continuous, remote, long-term monitoring and investigation of flow rates, growth, and other aspects of the vents via PAM.
To explore the potential of PAM, ONC deployed a hydrophone at MEF in 2018, and then upgraded the installation to a four-element array in 2023. Additionally, Dalhousie University’s Deep Acoustic Lander (Figure 4) was deployed and recovered in 2021 and 2023, further augmenting the time series (Smith and Barclay, 2023). Though still in its infancy, this study has already detected a large number of transient (i.e., of duration measurable in seconds or less), often impulsive, sounds characterizing the soundscape at MEF. These include chimney collapses, waterborne signals associated with earthquakes, and a number of other sounds whose origins are being investigated. A recent study reports that numerous such signals were captured by ONC’s hydrophones during the major seismic event of March 5–6, 2024. Through the investigation of power spectral density, ambient-noise coherence, and cross-correlation with other sensors at MEF, the same study highlighted other, longer-term changes in the MEF soundscape that may be associated with changes to the venting activity resulting from the increased seismicity in the region (Smith and Barclay, 2024).
Finally, PAM is also being explored as a tool for environmental impact assessment. Some marine organisms may use acoustic cues to select settlement locations around hydrothermal vents (Eggleston et al., 2016). Industrial activities, such as shipping and deep-sea mining, can potentially interfere with the local ecosystem by introducing changes in the soundscape, even though they may be located at significant distances (Chen et al., 2021). Understanding of the local soundscape relevant to the biological activity of a site is an important component of an effective environmental impact mitigation strategy (Lin et al., 2019).
Vent Biology
Numerous biological studies utilizing video imagery and samples collected from ROVs and submersibles have been conducted at Endeavour. They focused on describing the benthic assemblages inhabiting a range of hydrothermal vent conditions, from those on high-temperature black smoker chimneys to those sustained by broadly spread diffusive flows (Sarrazin et al., 1997; Tunnicliffe et al., 1997; Lelièvre et al., 2018; Murdock et al., 2021). The early studies of hydrothermal vent systems described a specialized fauna characterized by low species diversity, high biomass, and high levels of endemicity (i.e., species only occurring at vent environments; Tunnicliffe and Fowler, 1996; reviewed in Van Dover, 2000).
A key characteristic of typical vent fauna is successful associations between chemoautotrophic, symbiotic microorganisms and their macroinvertebrate hosts (Lonsdale, 1977; Corliss et al., 1979). Utilizing the chemical energy from sulfur, hydrogen, iron, and methane, vent microorganisms fix carbon not only in symbiont associations with host species but also as free-living cells or in extensive bacterial mats (Dick, 2019). Host-symbiont associations often achieve high densities and biomass surrounding the areas of hydrothermal fluid flow. At the Endeavour vents, the most conspicuous and abundant vent fauna assemblages are comprised of the siboglinid polychaete tubeworm Ridgeia piscesae, alvinelid polychaetes Paralvinella sulfincola (sulfide worm) and Paralvinella palmiiformis (palm worm), the limpet Lepetodrilus fucensis, and many other species of snails (Figure 5a-d, Sarrazin et al., 1997). Studies to date have inventoried close to 60 vent-associated species at Endeavour, with 12 endemic species not occurring anywhere else in the world (Fisheries and Oceans Canada, 2010). Sampling of macrofauna associated with tubeworm bushes near the Grotto edifice alone revealed up to 31 species occurring in substrate patches of less than 0.1 m2, and it highlighted the importance of keystone species such as R. piscesae in creating habitat complexity that enhances local biodiversity (Lelièvre et al., 2018).

FIGURE 5. A sample of habitat heterogeneity and biological diversity of vent-associated and vent periphery fauna at Endeavour. (a,b) Black smoker chimneys colonized by dense assemblages of R. piscesae tubeworms. (c,d) Typical assemblages that occur near diffusive hydrothermal flow, including alvinelid polychaetes (Paralvinella sulfincola, Paralvinella palmiiformis), polynoid scale worms (Branchinotogluma tunnicliffae), limpets (Lepetodrilus fucensis), and snails (Buccinum thermophilum). (e) Field of view of the Mothra vent field observatory camera showing the seafloor partially covered by white bacterial mats, Ridgeia piscesae tubeworms, Buccinum thermophilum gastropods, and the deep-sea spider crab, Macroregonia macrochira. (f) Vent periphery sulfide and (g,h) basalt structured seafloor that provide habitat for corals, sponges, and mobile macro- and megafauna. Jump to the flipbook version of this article for a slideshow of this figure. Image credits: ONC and CSSF–ROPOS. > High res figure
|
The roles of microbial diversity and production in controlling large-scale nutrient elemental cycling and ecosystem function have also been topics of studies based on the frequent sampling at Endeavour. Samples of diffusive sulfidic vent fluids helped to quantify microbial production pathways (denitrification, anammox, and dissimilatory nitrate reduction to ammonium), aiding global estimates of nitrogen (N) removal rates to the subsurface biosphere that represent 2.5%–3.5% of total marine N loss (Bourbonnais et al., 2012). Microbes were also the focus of a number of studies examining vent fauna host-symbiont relationships and population structure. The tubeworm Ridgeia piscesae, a keystone species, was found to have the same phylotype Gammaproteobacteria symbiont (Ca. Endorifitia persephone) as six other tubeworm species in the Eastern Pacific, revealing high levels of interconnectivity between the Northeast Pacific and the East Pacific Rise vents (Perez and Juniper, 2016). However, the same authors later uncovered multiple genotypes within E. persephone making up the symbiont assemblages of R. piscesae and argued that this genetic diversity could be an important predictor of resilience to environmental change (Perez and Juniper, 2017).
Since the installation of seafloor cables and platforms in the axial valley of the Endeavour Segment in 2010, in situ instruments and sensors, including time-lapse video imagery, have been providing new insights into the environmental controls over vent species community composition and biorhythms. At Main Endeavour Field, a video camera platform (TEMPO-Mini; Auffret et al., 2009), installed in collaboration with the French national institute for ocean science and technology (IFREMER), provided nearly 10 years of continuous data. The length of the video time series enabled analyses that, for the first time, established astronomical (tidal) and atmospheric (storm passages) forcing as a control on vent macrofauna behavior (Cuvelier et al., 2014, 2017; Lelièvre et al., 2017). The data revealed that mobile macrofauna, such as sea spiders (pycnogonids) and polychaete scale worms (polynoids), responded to the passage of winter storms 2.2 km above by regulating their biorhythms to the storm-triggered cyclical oscillations in the diffusive vent flow dynamics (Lelièvre et al., 2017). Video observations of picnogonids and scale worms living in association with R. piscesae tubeworm bushes that are supported by low-temperature diffuse venting also indicated that the animals respond to the currents generated by these storms. At the latitude of Endeavour, storm-induced currents have a four-day cycle due to the passage of the storms and a 16-hour cycle resulting from the inertial oscillations generated by the storm winds that can propagate to the seafloor as inertial internal waves. As the currents cyclically increase, they dilute the warm, low-oxygen vent fluids, and the animals can be observed moving deeper into the bush, disappearing from camera view. A study performed in waters 1,688 m deep at the EMSO-Azores Mid-Atlantic Ridge observatory (EMSO = European Multidisciplinary Seafloor and water column Observatory) corroborates these findings, as biological rhythms and circadian clock gene expression of the hydrothermal vent mussel Bathymodiolus azoricus were found to be directly tied to tidal cycles (Mat et al., 2020). Combined, these findings provide compelling evidence of more direct dynamical influences of the surface ocean and the planetary climate on deep ocean hydrothermal vent ecosystems than was previously thought. Furthermore, they highlight the importance of long-term observations supported by the NEPTUNE observatory in detecting faunal community changes at Endeavour in response to upper ocean climate variability.
A second seafloor camera installed at Mothra vent field in 2020 is further contributing to our understanding of the temporal dynamics of highly mobile and non-vent exclusive benthic megafauna, such as zoarcid and macrourid fishes and decapod crustaceans, by employing machine learning automatic classification and counting of the most abundant taxa (Carter, 2025; Figure 5e). NEPTUNE’s multiple video camera platforms, which cover a range of vent habitat types and incorporate embedded pipelines for automated imagery processing, can be used to inform MPA managers of long-term trends in faunal abundance and diversity (Aguzzi et al., 2020; Ortenzi et al., 2024).
A recent study focused on non-vent benthic megafauna inhabiting peripheral habitats (e.g., Figure 5f,g) located as much as a few kilometers away from the main active Endeavour vent sites. ROV video surveys conducted at Main Endeavour and High Rise vent fields revealed diverse assemblages dominated by slow growing sessile animals, such as rosselid vase sponges, alcyonacean corals, and crinoids (Neufeld et al., 2022). A key finding was that corals were nearly absent and rosselid sponges were found in very low abundances within 25–50 m of active chimneys but became progressively more abundant and diverse moving away from the vents; they occurred predominantly at bare basalt ridges and on walls of inactive sulfide chimneys. Species richness measured using rarefaction curves were significantly higher at inactive chimneys but never reached asymptotic values, demonstrating an undersampled and incomplete species catalogue (Neufeld et al., 2022). These results highlight the importance of studies that consider vent-periphery habitats covering a wider, landscape-scale habitat heterogeneity in order to uncover the true ecological “sphere of influence” (sensu Levin et al., 2016) and the true biodiversity conservation and MPA management value surrounding any hydrothermal vent system. Furthermore, parallel studies at Endeavour, such as Georgieva et al. (2020) that investigated microbiomes of vent-periphery sponges in the genus Spinularia, uncovered putative chemosynthetic Gammaproteobacteria (Thioglobaceae and Methylomonaceae) directly providing nutrition to the sponges, indicating that typical non-vent megafauna still benefit from symbiont associations deriving from dispersed vent fluids in the surroundings of active hydrothermal sites. Given deep-sea massive sulfide deposit mining activities being proposed for inactive vent sites around the globe (Jamieson and Gartman, 2020), the Endeavour Segment, which is protected under the TḥT MPA, therefore becomes a natural study and monitoring site for further exploration of the importance of vent-periphery habitats, biodiversity, and resilience to human impacts.
Photogrammetric mosaics produced by repeated flyovers using remote or autonomously operated vehicles with dedicated camera systems (Van Audenhaege et al., 2024) allow monitoring of ecological dynamics from vent-edifice to centimeter scales. Motion photogrammetry has been used to generate highly accurate habitat terrain models, with high predictive power for faunal assemblage distribution (Gerdes et al., 2019). The most important community structuring variables in these habitat models are often distances to diffuse and black fluid exits, as well as the height of the chimney complex (Gerdes et al., 2019; Girard et al., 2020). At Endeavour, regular maintenance visits to the observatory with a scientific ROV enabled assembly of a sequence of 3D photogrammetry models from repeat visits to the Mothra vent field. Data are being analyzed, with preliminary results providing insights into how chimney accretion and erosion affect spatial distribution and community succession of vent fauna (Tom Kwasnitschka, GEOMAR Helmholtz Centre for Ocean Research Kiel, pers. comm., October 22, 2024).
Additionally, ROV survey data (video and navigation) collected during observatory maintenance expeditions can be mined to produce kernel density “heat maps” of significant ecosystem components, indicators, and stressors (Juniper et al., 2019). As the observatory maintenance and operations occur on a yearly basis, these maps are continuously updated and are used as essential MPA spatial management tools by quantitatively assessing research pressure on the vents (e.g., ROV tracks and sampling efforts), hotspots of biodiversity associated with vents (typical chemosynthetic communities), and vent periphery habitats (e.g., corals and sponges) (Fisheries and Oceans Canada, 2025). An interactive map is available online with multiple geographical information system (GIS) layers of all observatory maintenance activities, spatial distribution of venting habitat structures (e.g., active and inactive edifices), and associated biodiversity.
Conclusion
Since deployment of scientific instrumentation at the Endeavour MPA in the fall of 2010, a total of 103 peer-reviewed papers have been published, as well as 10 dissertations and one book chapter (in The Sound of Hydrothermal Vents, Smith and Barclay, 2023). Of the 103 journal articles, 51 were based directly on sensor data archived in Oceans 3.0 and/or discrete samples collected on maintenance expeditions, while 28 articles used ONC data along with data from other sources (e.g., earthquake, acoustics). The remaining 24 articles were either review/overview articles or articles supported by research enabled by ONC. The internet access and the power offered by the deep-sea observatory promoted the development of new hydrothermal vent and seafloor monitoring technology, while the research resulted in significant advancement in our understanding of the geophysics, vent biology, and oceanography of the MPA.
The observations collected by cabled sensors and during the repeated visits for infrastructure maintenance are freely available (unless there is a known student thesis that could be affected by early release of the data). There is growing understanding of hydrothermal ecosystems’ functions, their connectivity to other ecosystems, their benefit to humanity, and their role in the ocean’s chemical balance. However, a clear picture of the “value” of hydrothermal sites to weigh against disturbance, for example by deep-sea mining or scientific sampling, is not complete (Turner et al., 2019). The objective of continuous, long time-series monitoring of vent sites by seafloor observatories is to enable observation of natural disturbances and how succession proceeds afterwards to understand what level of anthropogenic disturbance or scientific sampling might be tolerated. In addition, real-time monitoring is essential for monitoring episodic natural perturbations. A framework is being developed among the Canadian government and scientific institutions for a rapid response to a perturbation of the hydrothermal system at the Endeavour Segment to gain observations of the changes in the water column chemistry and the benthic and pelagic ecosystems.
The Endeavour MPA is remote, 300 km offshore and in 2.2 km of water, and seemingly clear of threats that impact coastal oceans. However, it is not isolated from ocean acidification, microplastic pollution, hypoxia, and even storm intensity. Real-time data and regular, yearly maintenance visits to Endeavour monitor change due to natural processes, pollution, and climate change.
With the Endeavour MPA subsumed into the much larger offshore TḥT MPA (Figure 6), now encompassing multiple ecologically or biologically significant marine areas, remote monitoring strategies are expected to change in accordance with new MPA conservation and management goals. While the cabled seafloor sensors will play a crucial role, other complementary monitoring and infrastructure upgrades are needed to continue increasing scientific understanding, to contribute to improved management or conservation, and to monitor the effectiveness of the new MPA protections.

FIGURE 6. Map showing the boundaries (yellow polygon) of the offshore TḥT MPA. Boundaries encompass all known hydrothermal vent fields within Canada, at least 47 seamounts, and hundreds of seamount-like features. The ONC observatories are delineated by orange squares. Image credit: ONC. > High res figure
|
Acknowledgments
ONC is primarily funded by the Canada Foundation for Innovation, Government of Canada, University of Victoria, and Government of British Columbia. Thank you to Mark Rankin for preparing Figure 1, and to Norman Coloma for Figure 4. Many thanks to the captains and crew of R/V T.G. Thompson, CCGS John P. Tully, and E/V Nautilus, as well as the crews of the remotely operated vehicles ROPOS, Hercules, Odysseus, and Millennium. Finally, and most importantly, we would like to thank all the researchers who have been fascinated by the Endeavour Segment and provided the impetus for its designation as an MPA. In particular, we thank S.K. Juniper (in memoriam) for his passion, enthusiasm, and curiosity, all of which spurred a multitude of new and exciting discoveries about Endeavour vent communities.

