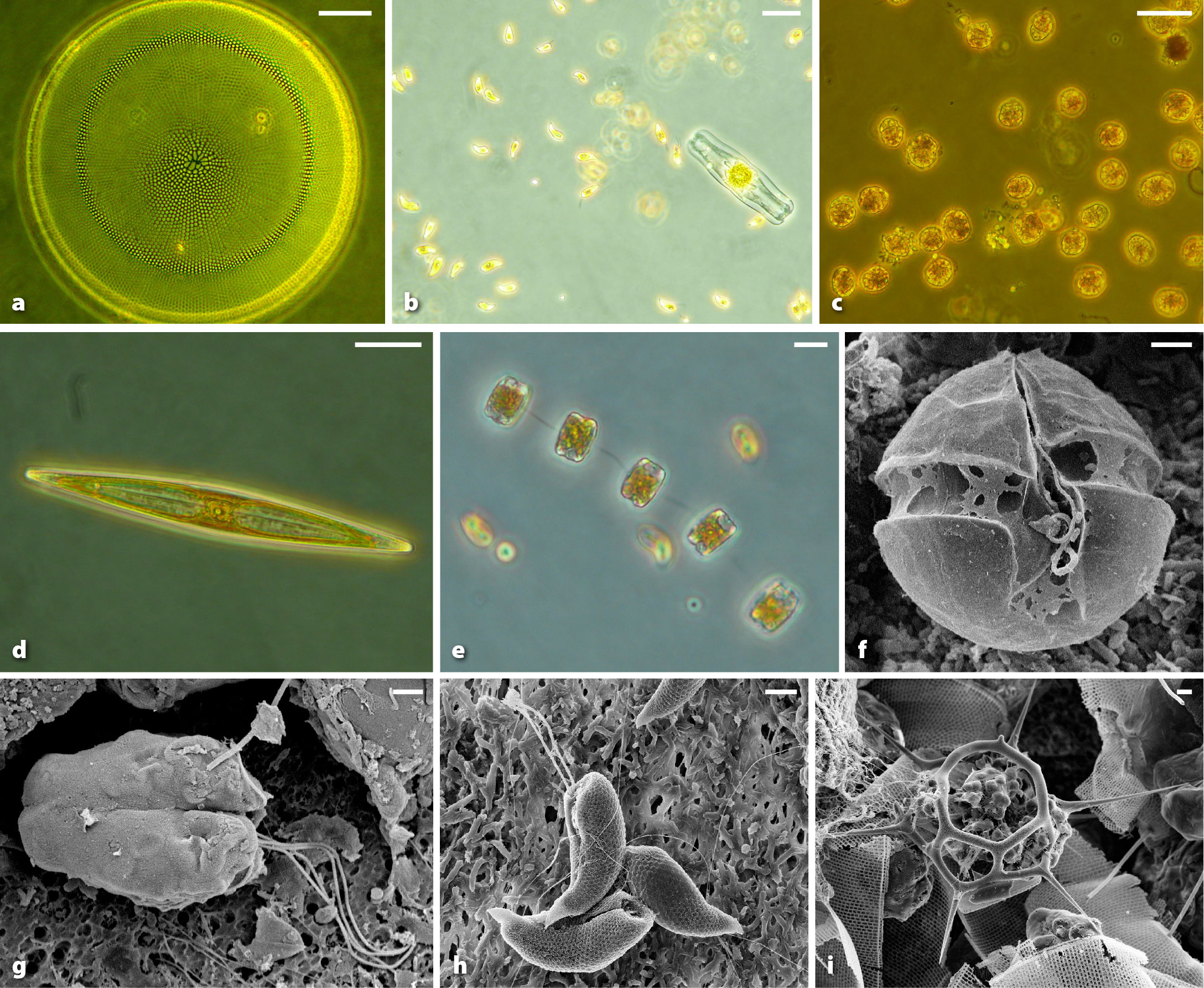
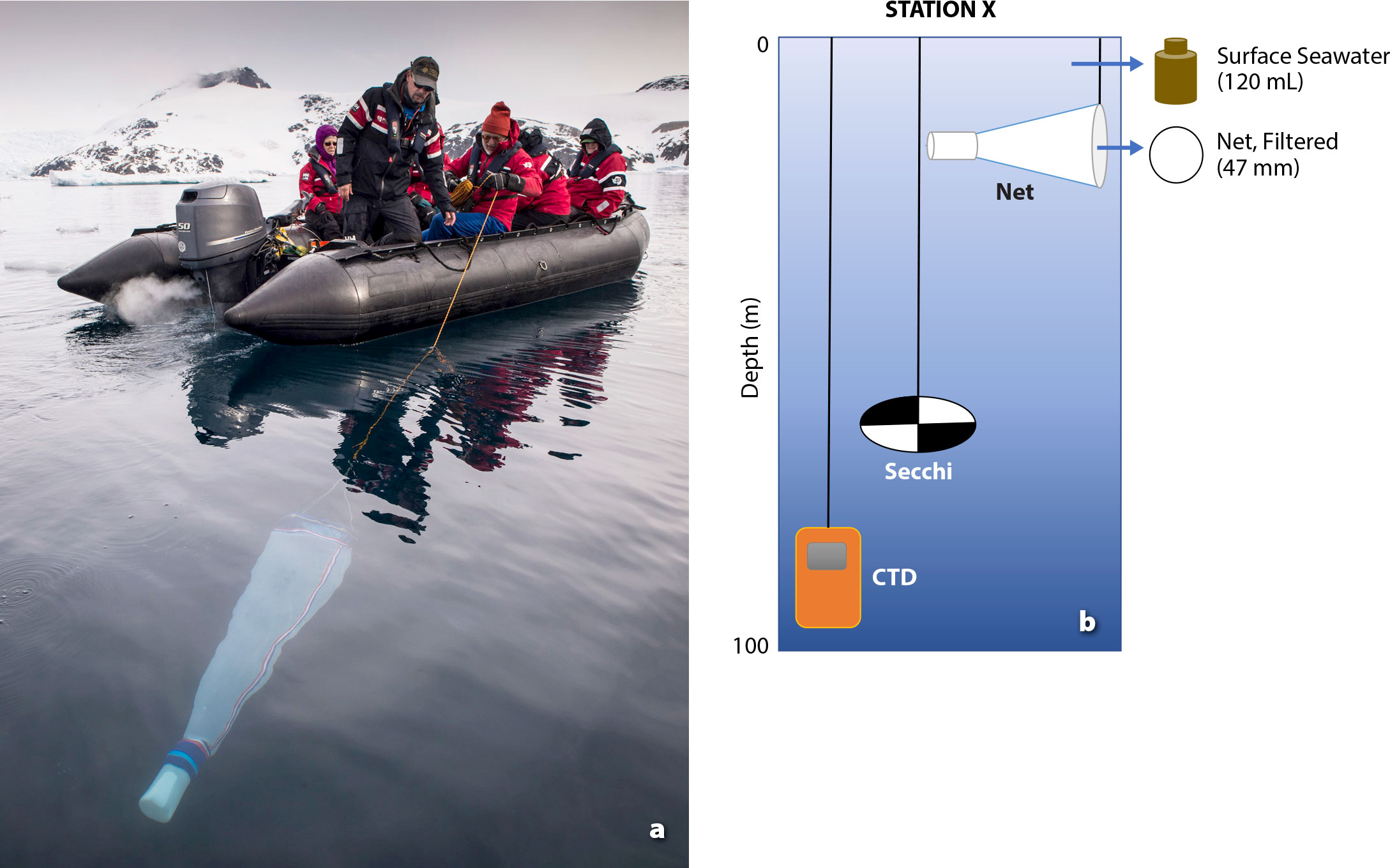
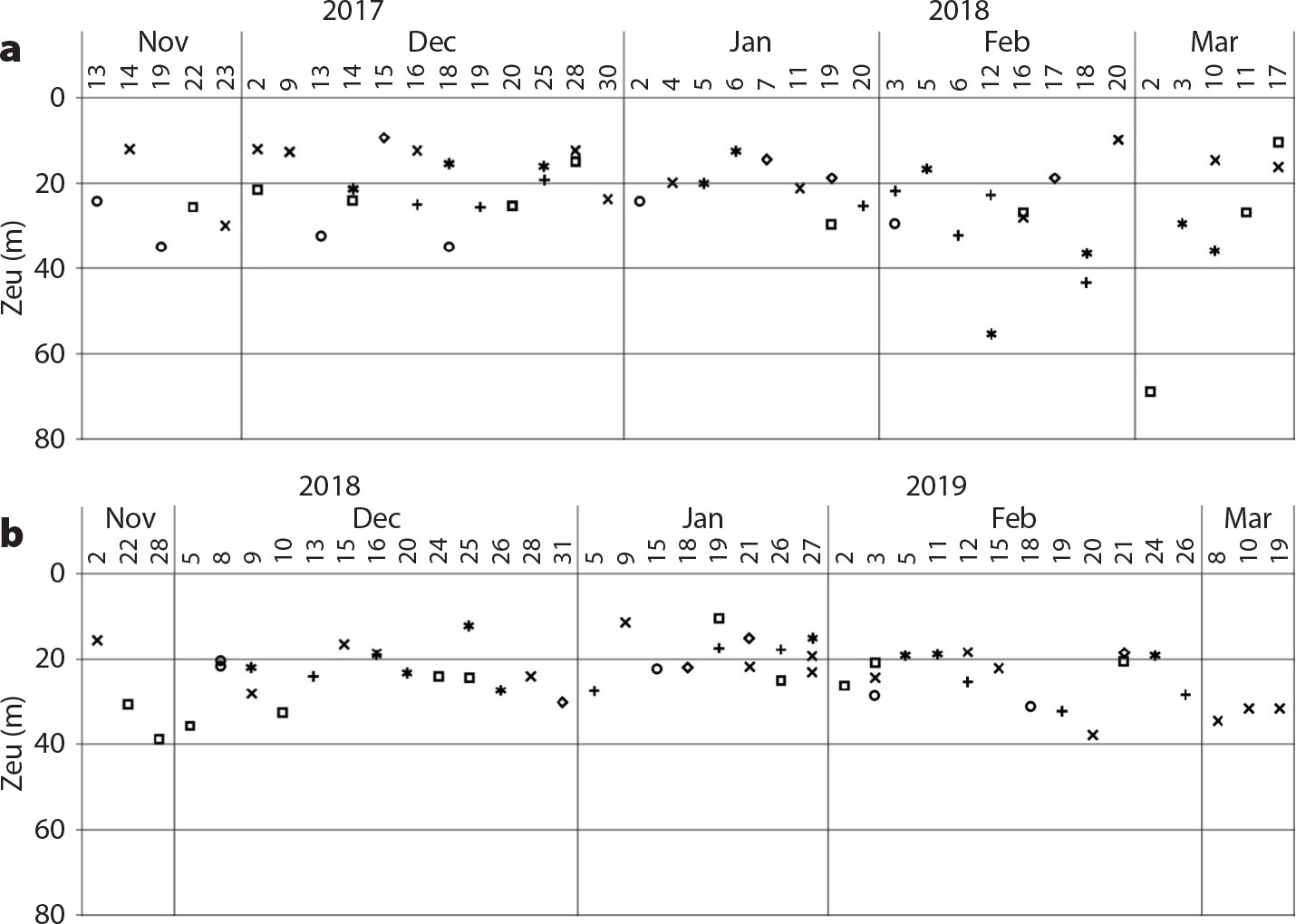
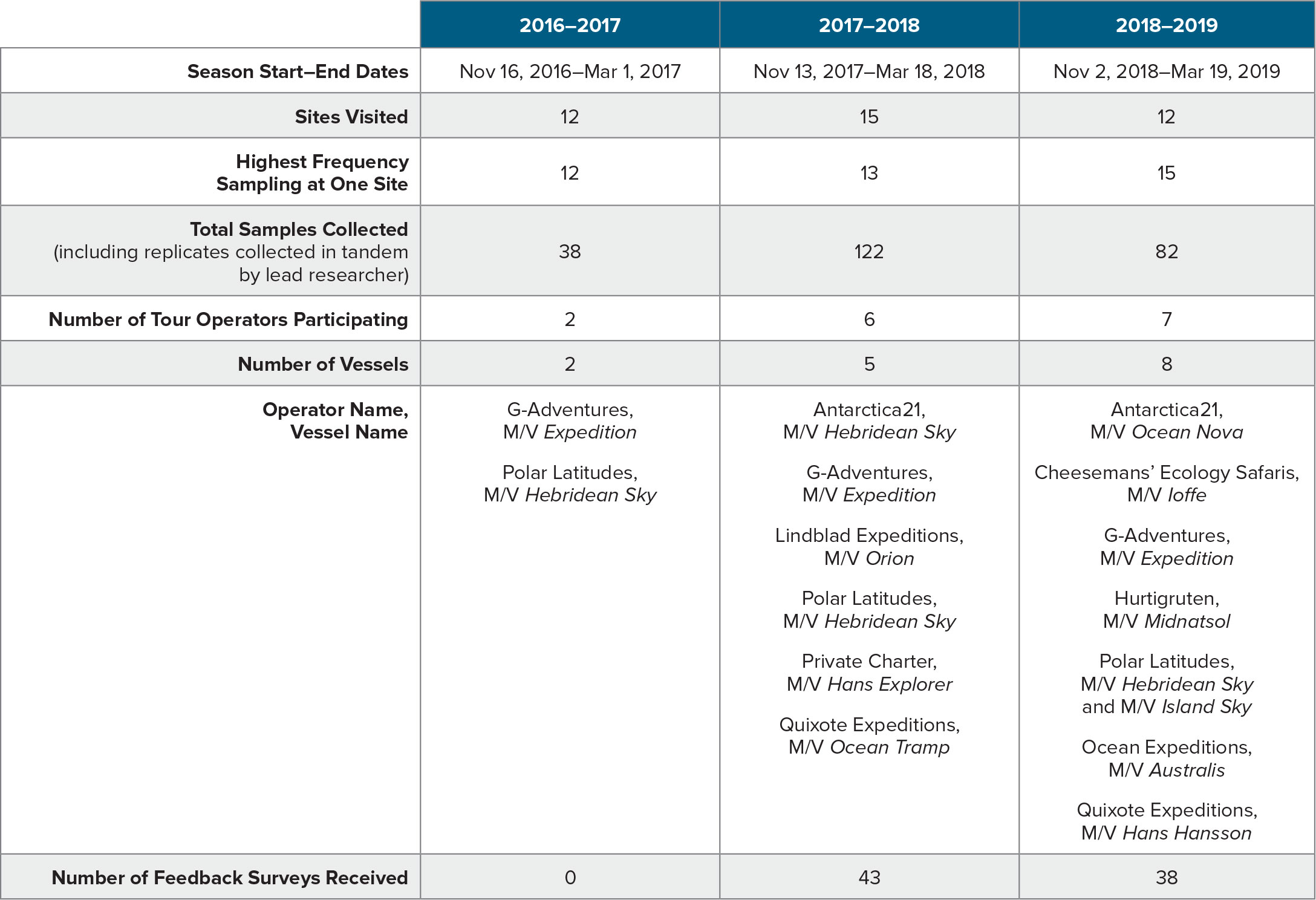
Aas, E., J. Høkedal, and K. Sørensen. 2014. Secchi depth in the Oslofjord-Skagerrak area: Theory, experiments and relationships to other quantities. Ocean Science 10:177–199, https://doi.org/10.5194/os-10-177-2014.
Bender, N.A., K. Crosbie, and H.J. Lynch. 2016. Patterns of tourism in the Antarctic Peninsula region: A 20-year analysis. Antarctic Science 28(03):194–203, https://doi.org/10.1017/S0954102016000031.
Bernard, K.S., and D.K. Steinberg. 2013. Krill biomass and aggregation structure in relation to tidal cycle in a penguin foraging region off the Western Antarctic Peninsula. ICES Journal of Marine Science 70(4):834–849, https://doi.org/10.1093/icesjms/fst088.
Bonney, R., C.B. Cooper, J. Dickinson, S. Kelling, T. Phillips, K.V. Rosenberg, and J. Shirk. 2009. Citizen science: A developing tool for expanding science knowledge and scientific literacy. BioScience 59(11):977–984, https://doi.org/10.1525/bio.2009.59.11.9.
Brandini, F.P., and J. Rebello. 1994. Wind field effect on hydrography and chlorophyll dynamics in the coastal pelagial of Admiralty Bay, King George Island, Antarctica. Antarctic Science 6(4):433-442, https://doi.org/10.1017/S0954102094000672.
Constable, A.J., J. Melbourne-Thomas, S.P. Corney, K.R. Arrigo, C. Barbraud, D.K. Barnes, N.L. Bindoff, P.W. Boyd, A. Brandt, D.P. Costa, and A.T. Davidson. 2014. Climate change and Southern Ocean ecosystems I: How changes in physical habitats directly affect marine biota. Global Change Biology 20(10):3,004–3,025, https://doi.org/10.1111/gcb.12623.
Cook, A.J., P.R. Holland, M.P. Meredith, T. Murray, A. Luckman, and D.G. Vaughan. 2016. Ocean forcing of glacier retreat in the Western Antarctic Peninsula. Science 353(6296):283–286, https://doi.org/10.1126/science.aae0017.
Corbisier, T.N., M.A. Petti, R.S. Skowronski, and T.A. Brito. 2004. Trophic relationships in the nearshore zone of Martel Inlet (King George Island, Antarctica): δ13C stable-isotope analysis. Polar Biology 27(2):75–82, https://doi.org/10.1007/s00300-003-0567-z.
Crall, A.W., G.J. Newman, T.J. Stohlgren, K.A. Holfelder, J. Graham, and D.M. Waller. 2011. Assessing citizen science data quality: An invasive species case study. Conservation Letters 4(6):433–442, https://doi.org/10.1111/j.1755-263X.2011.00196.x.
Cusick, A.M. 2017a. Leveraging citizen science to understand glacial meltwater impacts on phytoplankton communities within fjords of the Western Antarctic Peninsula. Poster presented at the Gordon Research Conference on Polar Marine Science, March 26–31, 2017, Ventura, California.
Cusick, A.M. 2017b. The invisible forest: Using citizen science to improve understanding of Antarctic phytoplankton. Paper presented at the AECO-IAATO Polar Field Staff Conference, October 1–4, 2017, Hveragerði, Iceland.
Cusick, A.M. 2018a. FjordPhyto Guest Scientist A21 2017 w music. YouTube video. December 18, 2018, https://youtu.be/n1yKoFHni_U.
Cusick, A.M. 2018b. Antarctic ambassadors learning to love fjord phytoplankton. Online presentation at the Association for Polar Early Career Scientists (APECS) 2018 Conference, May 16, 2018.
Cusick, A.M. 2018c. FjordPhyto: Understanding Antarctic phytoplankton through citizen science. Paper presented at the Scientific Committee on Antarctic Research (SCAR) and the International Arctic Science Committee (IASC) Polar 2018 Conference, June 19–23, 2018, Davos, Switzerland.
Cusick, A.M. 2019. Citizen Scientists in Antarctica a new approach to understand climate change affected environments with FjordPhyto. Paper presented at CitSci2019 Conference, March 13–17, 2019, Raleigh, North Carolina.
De Lima, D.T., G.O. Moser, F.R. Piedras, L.C. da Cunha, D.R. Tenenbaum, M.B. Tenório, and J.J. Barrera-Alba. 2019. Abiotic changes driving microphytoplankton functional diversity in Admiralty Bay, King George Island (Antarctica). Frontiers in Marine Science 6:638, https://doi.org/10.3389/fmars.2019.00638.
Dierssen, H.M., R.C. Smith, and M. Vernet. 2002. Glacial meltwater dynamics in coastal waters west of the Antarctic peninsula. Proceedings of the National Academy of Sciences of the United States of America 99(4):1,790–1,795, https://doi.org/10.1073/pnas.032206999.
Edelson, D.C., S.L. Kirn, and Workshop Participants. 2018. Designing citizen science for both science and education: A workshop report. Technical Report No. 2018-01, BSCS Science Learning, Colorado Springs, CO.
Edler, L., and M. Elbrächter. 2010. The Utermöhl method for quantitative phytoplankton analysis. Pp. 13–20 in Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis. B. Karlson, C. Cusack and E. Bresnan, eds, Intergovernmental Oceanographic Commission of UNESCO. IOC Manuals and Guides, no. 55, UNESCO, Paris.
Espinasse, B., M. Zhou, Y. Zhu, E.L. Hazen, A.S. Friedlaender, D.P. Nowacek, D. Chu, and F. Carlotti. 2012. Austral fall-winter transition of mesozooplankton assemblages and krill aggregations in an embayment west of the Antarctic Peninsula. Marine Ecology Progress Series 452:63–80, https://doi.org/10.3354/meps09626.
Falkowski, P.G. 2002. The ocean’s invisible forest. Scientific American 287(2):54–61, https://doi.org/10.1038/scientificamerican0802-54.
Garibotti, I.A., M. Vernet, and M.E. Ferrario. 2005. Annually recurrent phytoplanktonic assemblages during summer in the seasonal ice zone west of the Antarctic Peninsula (Southern Ocean). Deep Sea Research Part I 52(10):1,823–1,841, https://doi.org/10.1016/j.dsr.2005.05.003.
Grange, L.J., and C.R. Smith. 2013. Megafaunal communities in rapidly warming fjords along the West Antarctic Peninsula: Hotspots of abundance and beta diversity. PLOS ONE 8(12):e77917, https://doi.org/10.1371/journal.pone.0077917.
Haberman, K.L., R.M. Ross, and L.B. Quetin. 2003. Diet of the Antarctic krill (Euphausia superba Dana): II. Selective grazing in mixed phytoplankton assemblages. Journal of Experimental Marine Biology and Ecology 283(1–2):97–113, https://doi.org/10.1016/S0022-0981(02)00467-7.
Henley, S.F., O.M. Schofield, K.R. Hendry, I.R. Schloss, D.K. Steinberg, C. Moffat, L.S. Peck, D.P. Costa, D.C. Bakker, C. Hughes, and P.D. Rozema. 2019. Variability and change in the west Antarctic Peninsula marine system: Research priorities and opportunities. Progress in Oceanography 173:208–237, https://doi.org/10.1016/j.pocean.2019.03.003.
Hernando, M., I.R. Schloss, G. Malanga, G.O. Almandoz, G.A. Ferreyra, M.B. Aguiar, and S. Puntarulo. 2015. Effects of salinity changes on coastal Antarctic phytoplankton physiology and assemblage composition. Journal of Experimental Marine Biology and Ecology 466:110–119, https://doi.org/10.1016/j.jembe.2015.02.012.
Hou, W., Z. Lee, and A.D. Weidemann. 2007. Why does the Secchi disk disappear? An imaging perspective. Optics Express 15(6):2,791–2,802, https://doi.org/10.1364/OE.15.002791.
IAATO (International Association of Antarctica Tour Operators). 2009. Report of the International Association of Antarctica Tour Operators 2008–2009 Under Article III (2) of the Antarctic Treaty. ATCM32 IP33, Antarctic Treaty Consultative Meeting, Baltimore, USA, https://iaato.org/past-iaato-information-papers.
IAATO. 2019a. IAATO Overview of Antarctic Tourism: 2018–19 Season and Preliminary Estimates for 2019–20 Season. ATCM 17, IP140, Antarctic Treaty Consultative Meeting, Prague, Czech Republic, https://iaato.org/current-iaato-information-papers.
IAATO. 2019b. What is IAATO’s Mission? https://iaato.org/frequently-asked-questions#what-is-iaatos-mission.
Kennicutt, M.C., S.L. Chown, J.J. Cassano, D. Liggett, R. Massom, L.S. Peck, S.R. Rintoul, J.W. Storey, D.G. Vaughan, T.J. Wilson, and W.J. Sutherland. 2014. Six priorities for Antarctic science. Nature 512(7512):23–25, https://doi.org/10.1038/512023a.
Kim, H., H.W. Ducklow, D. Abele, E.M. Ruiz Barlett, A.G. Buma, M.P. Meredith, P.D. Rozema, O.M. Schofield, H.J. Venables, and I.R. Schloss. 2018. Inter-decadal variability of phytoplankton biomass along the coastal West Antarctic Peninsula. Philosophical Transactions of the Royal Society A 376(2122):20170174, https://doi.org/10.1098/rsta.2017.0174.
Kozlowski, W., M. Vernet, and S.K. Lamerdin. 1995. Predominance of cryptomonads and diatoms in Antarctic coastal waters. Antarctic Journal of the United States 30:267–268.
Lange, P.K., D.R. Tenenbaum, V.M. Tavano, R. Paranhos, and L.D.S. Campos. 2015. Shifts in microphytoplankton species and cell size at Admiralty Bay, Antarctica. Antarctic Science 27(3):225–239, https://doi.org/10.1017/S0954102014000571.
Larsen, J.N., O.A. Anisimov, A. Constable, A.B. Hollowed, N. Maynard, P. Prestrud, T.D. Prowse, and J.M.R. Stone. 2014. Polar regions. Pp. 1,567–1,612 in Climate Change 2014: Impacts, Adaptation, and Vulnerability: Part B. Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. V.R. Barros, C.B. Field, D.J. Dokken, M.D. Mastrandrea, K.J. Mach, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L. White, eds, Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA.
Lee Cusick, A. 2017. “FjordPhyto: Antarctic Citizen Science Project.” UC San Diego: Center for Marine Biodiversity and Conservation, https://escholarship.org/uc/item/9m05m5g3.
Lin, Y., N. Cassar, A. Marchetti, C. Moreno, H. Ducklow, and Z. Li. 2017. Specific eukaryotic plankton are good predictors of net community production in the Western Antarctic Peninsula. Scientific Reports 7(1):14845, https://doi.org/10.1038/s41598-017-14109-1.
Mascioni, M. 2019. Antarctic tourism as a research tool: Citizen Science monitoring phytoplankton community in the Western Antarctic Peninsula (WAP). Paper presented at the Scientific Committee on Antarctic Research’s (SCAR) Standing Committee on Antarctic Humanities and Social Sciences (SC-HASS) April 3–5, 2019, Ushuaia, Argentina.
Mascioni, M., G.O. Almandoz, A.O. Cefarelli, A. Cusick, M.E. Ferrario, and M. Vernet. 2019. Phytoplankton composition and bloom formation in unexplored nearshore waters of the western Antarctic Peninsula. Polar Biology 42:1,859–1,872, https://doi.org/10.1007/s00300-019-02564-7.
Magnuson, J.J. 1990. Long-term ecological research and the invisible present. BioScience 40(7):495–501, https://doi.org/10.2307/1311317.
McKinley, D.C., A.J. Miller-Rushing, H.L. Ballard, R. Bonney, H. Brown, S.C. Cook-Patton, D.M. Evans, R.A. French, J.K. Parrish, T.B. Phillips, and S.F. Ryan. 2017. Citizen science can improve conservation science, natural resource management, and environmental protection. Biological Conservation 208:15–28, https://doi.org/10.1016/j.biocon.2016.05.015.
Mendes, C.R.B., V.M. Tavano, M.C. Leal, M.S. de Souza, V. Brotas, and C.A.E. Garcia. 2013. Shifts in the dominance between diatoms and cryptophytes during three late summers in the Bransfield Strait (Antarctic Peninsula). Polar Biology 36(4):537–547, https://doi.org/10.1007/s00300-012-1282-4.
Meredith, M.P., H.J. Venables, A. Clarke, H.W. Ducklow, M. Erickson, M.J. Leng, J.M. Lenaerts, and M.R. van den Broeke. 2013. The freshwater system west of the Antarctic Peninsula: Spatial and temporal changes. Journal of Climate 26(5):1,669–1,684, https://doi.org/10.1175/JCLI-D-12-00246.1.
Miller, J.D. 2016. Civic Scientific Literacy in the United States in 2016. A report prepared for the National Aeronautics and Space Administration by the University of Michigan, Ann Arbor, MI.
Miller-Rushing, A., R. Primack, and R. Bonney. 2012. The history of public participation in ecological research. Frontiers in Ecology and the Environment 10(6):285–290, https://doi.org/10.1890/110278.
Mitchell, B.G., and O. Holm-Hansen. 1991. Observations of modeling of the Antarctic phytoplankton crop in relation to mixing depth. Deep Sea Research Part A 38(8–9):981–1,007, https://doi.org/10.1016/0198-0149(91)90093-U.
Moline, M.A., H. Claustre, T.K. Frazer, O. Schofield, and M. Vernet. 2004. Alteration of the food web along the Antarctic Peninsula in response to a regional warming trend. Global Change Biology 10(12):1,973–1,980, https://doi.org/10.1111/j.1365-2486.2004.00825.x.
Nicol, S., A. Bowie, S. Jarman, D. Lannuzel, K.M. Meiners, and P. Van Der Merwe. 2010. Southern Ocean iron fertilization by baleen whales and Antarctic krill. Fish and Fisheries 11(2):203–209, https://doi.org/10.1111/j.1467-2979.2010.00356.x.
Nowacek, D.P., A.S. Friedlaender, P.N. Halpin, E.L. Hazen, D.W. Johnston, A.J. Read, B. Espinasse, M. Zhou, and Y. Zhu. 2011. Super-aggregations of krill and humpback whales in Wilhelmina Bay, Antarctic Peninsula. PLOS ONE 6(4):e19173, https://doi.org/10.1371/journal.pone.0019173.
Pan, B.J., M. Vernet, R.A. Reynolds, and B.G. Mitchell. 2019. The optical and biological properties of glacial meltwater in an Antarctic fjord. PLOS ONE 14(2):e0211107, https://doi.org/10.1371/journal.pone.0211107.
Phillips, T., M. Ferguson, M. Minarchek, N. Porticella, and R. Bonney. 2014. User’s Guide for Evaluating Learning Outcomes in Citizen Science. Cornell Lab of Ornithology, Ithaca, NY, https://www.birds.cornell.edu/citizenscience/wp-content/uploads/2018/10/USERS-GUIDE_linked.pdf.
Rodriguez, F., M. Varela, and M. Zapata. 2002. Phytoplankton assemblages in the Gerlache and Bransfield Straits (Antarctic Peninsula) determined by light microscopy and CHEMTAX analysis of HPLC pigment data. Deep Sea Research Part II 49(4–5):723–747, https://doi.org/10.1016/S0967-0645(01)00121-7.
Ross, R.M., E.E. Hofmann, and L.B. Quetin, eds. 1996. Foundations for Ecological Research West of the Antarctic Peninsula, vol. 70. American Geophysical Union, Washington, DC, https://doi.org/10.1029/AR070.
Ross, R.M., L.B. Quetin, K.S. Baker, M. Vernet, and R.C. Smith. 2000. Growth limitation in young Euphausia superba under field conditions. Limnology and Oceanography 45:31–43, https://doi.org/10.4319/lo.2000.45.1.0031.
Rozema, P.D., H.J. Venables, W.H. Van De Poll, A. Clarke, M.P. Meredith, and A.G.J. Buma. 2017. Interannual variability in phytoplankton biomass and species composition in northern Marguerite Bay (West Antarctic Peninsula) is governed by both winter sea ice cover and summer stratification. Limnology and Oceanography 62(1):235–252, https://doi.org/10.1002/lno.10391.
Saba, G.K., W.R. Fraser, V.S. Saba, R.A. Iannuzzi, K.E. Coleman, S.C. Doney, H.W. Ducklow, D.G. Martinson, T.N. Miles, D.L. Patterson-Fraser, and others. 2014. Winter and spring controls on the summer food web of the coastal West Antarctic Peninsula. Nature Communications 5, 4318, https://doi.org/10.1038/ncomms5318.
Sailley, S.F., H.W. Ducklow, H.V. Moeller, W.R. Fraser, O.M. Schofield, D.K. Steinberg, L.M. Garzio, and S.C. Doney. 2013. Carbon fluxes and pelagic ecosystem dynamics near two western Antarctic Peninsula Adélie penguin colonies: An inverse model approach. Marine Ecology Progress Series 492:253–272, https://doi.org/10.3354/meps10534.
Santora, J.A., and C.S. Reiss. 2011. Geospatial variability of krill and top predators within an Antarctic submarine canyon system. Marine Biology 158(11):2,527–2,540, https://doi.org/10.1007/s00227-011-1753-0.
Sardet, C. 2015. Plankton: Wonders of the Drifting World. University of Chicago Press, https://doi.org/10.7208/chicago/9780226265346.001.0001.
Schloss, I.R., D. Abele, S. Moreau, S. Demers, A.V. Bers, O. González, and G.A. Ferreyra. 2012. Response of phytoplankton dynamics to 19-year (1991–2009) climate trends in Potter Cove (Antarctica). Journal of Marine Systems 92(1):53–66, https://doi.org/10.1016/j.jmarsys.2011.10.006.
Schofield, O., G. Saba, K. Coleman, F. Carvalho, N. Couto, H. Ducklow, Z. Finkel, A. Irwin, A. Kahl, T. Miles, and M. Montes-Hugo. 2017. Decadal variability in coastal phytoplankton community composition in a changing West Antarctic Peninsula. Deep Sea Research Part I 124:42–54, https://doi.org/10.1016/j.dsr.2017.04.014.
Sear, D. 2018. Plankton BioBlitz | Antarctica | Lindblad Expeditions-National Geographic. YouTube video, January 8, 2018, https://www.youtube.com/watch?v=ORrVYzM6da0.
Shirk, J., H. Ballard, C. Wilderman, T. Phillips, A. Wiggins, R. Jordan, E. McCallie, M. Minarchek, B. Lewenstein, M. Krasny, and R. Bonney. 2012. Public participation in scientific research: A framework for deliberate design. Ecology and Society 17(2):29, https://doi.org/10.5751/ES-04705-170229.
Smith, C.R., S. Mincks, and D.J. DeMaster. 2008. The FOODBANCS project: Introduction and sinking fluxes of organic carbon, chlorophyll-a and phytodetritus on the Western Antarctic Peninsula continental shelf. Deep Sea Research Part II 55(22):2,404–2,414, https://doi.org/10.1016/j.dsr2.2008.06.001.
Thomson, R.E., and I.V. Fine. 2003. Estimating mixed layer depth from oceanic profile data. Journal of Atmospheric and Oceanic Technology 20(2):319–329, https://doi.org/10.1175/1520-0426(2003)020<0319:EMLDFO>2.0.CO;2.
Turner, J., H. Lu, I. White, J.C. King, T. Phillips, J.S. Hosking, T.J. Bracegirdle, G.J. Marshall, R. Mulvaney, and P. Deb. 2016. Absence of 21st century warming on Antarctic Peninsula consistent with natural variability. Nature 535(7612):411, https://doi.org/10.1038/nature18645.
US Federal Government. 2019. The Federal Crowdsourcing and Citizen Science Toolkit, https://www.citizenscience.gov/toolkit/#.
Varela, M., E. Fernandez, and P. Serret. 2002. Size-fractionated phytoplankton biomass and primary production in the Gerlache and south Bransfield Straits (Antarctic Peninsula) in Austral summer 1995–1996. Deep Sea Research Part II 49(4-5):749–768, https://doi.org/10.1016/S0967-0645(01)00122-9.
Vernet, M., D. Martinson, R. Iannuzzi, S. Stammerjohn, W. Kozlowski, K. Sines, R. Smith, and I. Garibotti. 2008. Primary production within the sea-ice zone west of the Antarctic Peninsula: I—Sea ice, summer mixed layer, and irradiance. Deep Sea Research Part II 55(18–19):2,068–2,085, https://doi.org/10.1016/j.dsr2.2008.05.021.
Walsh, N. 2019. See the world and make a difference on a ‘Citizen Science’ Expedition. The New York Times, https://www.nytimes.com/2019/03/27/travel/see-the-world-and-make-a-difference-on-a-citizen-science-expedition.html.
Wernand, M.R. 2010. On the history of the Secchi disc. Journal of the European Optical Society-Rapid Publications, vol. 5, https://doi.org/10.2971/jeos.2010.10013s.
Wiggins, A., G. Newman, R.D. Stevenson, and K. Crowston. 2011. Mechanisms for data quality and validation in citizen science. Pp. 14–19 in 2011 IEEE Seventh International Conference on e-Science Workshops, December 5–8, 2011, Stockholm, Sweden, IEEE, https://doi.org/10.1109/eScienceW.2011.27.